ņä£ ļĪĀ
ņ£ä ņĪ░ņ¦ü ļ░░ņ¢æņØä ĒåĄĒĢ£ ĒŚ¼ļ”¼ņĮöļ░ĢĒä░ ĒīīņØ╝ļĪ£ļ”¼(Helicobacter pylori)ĻĘĀņØś Ļ▓ĆņČ£ņØĆ ņŗżļĪ£ Ēśüļ¬ģņĀüņØĖ ļ░£Ļ▓¼ņØ┤ļŗż. ĻĖ░ņĪ┤ņØś pH 2ļéś 3 ņĀĢļÅäņØĖ ņĢäņŻ╝ Ļ░ĢļĀźĒĢ£ ņ£äņé░ ĒÖśĻ▓ĮņŚÉņä£ļŖö ņ¢┤ļŖÉ ĻĘĀļÅä ĻĄ░ļØĮĒÖöĒĢĀ ņłś ņŚåņØä Ļ▓āņØ┤ļØ╝ļŖö ņØ╝ļ░śņĀüņØĖ ļ»┐ņØīņŚÉļÅä ļČłĻĄ¼ĒĢśĻ│Ā ĒŚ¼ļ”¼ņĮöļ░ĢĒä░ ĒīīņØ╝ļĪ£ļ”¼ ļ░░ņ¢æņØś ņä▒Ļ│ĄĻ│╝ ĒĢ©Ļ╗ś ļ░░ņ¢æļÉ£ ĻĘĀņŻ╝Ļ░Ć ņ£ä, ņŗŁņØ┤ņ¦Ćņן ĻČżņ¢æņØ┤ļéś ņ£äņŚ╝Ļ│╝ Ļ░ÖņØĆ ņ£ä ņ¦łĒÖśĻ│╝ņØś ņŚ░Ļ┤Ćņä▒ņØä ĻĘ£ļ¬ģĒĢ£ ņŚģņĀüņ£╝ļĪ£ ĒśĖņŻ╝ņØś Marshall ļ░Ģņé¼ņÖĆ Warren ĻĄÉņłśļŖö 2005ļģä ļģĖļ▓© ņØśĒĢÖņāØļ”¼ĒĢÖņāüņØä ļ░øĻ▓ī ļÉśņŚłļŗż[1-3]. ņØ┤ļ¤¼ĒĢ£ ņä▒Ļ│╝ļŖö ņ×äņāüņŚÉņä£ ņ£ä ļ░Å ņŗŁņØ┤ņ¦Ćņן ĻČżņ¢æņØś ļ░£ņāØņØĆ ļ¼╝ļĪĀ ņ×¼ļ░£Ļ│╝ ĒĢ®ļ│æņ”ØņØś ļ░®ņ¦Ćļź╝ ņ£äĒĢ┤ņä£ļŖö ļ░śļō£ņŗ£ ĒŚ¼ļ”¼ņĮöļ░ĢĒä░ ĒīīņØ╝ļĪ£ļ”¼ļź╝ ņĀ£ĻĘĀņŗ£ņ╝£ņĢ╝ļ¦ī ĒĢ£ļŗżļŖö ņé¼ņŗżļĪ£ ņØ┤ņ¢┤ņĀĖ, ĻĖ░ņĪ┤ņØś ŌĆ£No acid, No ulcerŌĆØļØ╝ļŖö ņåīĒÖöņä▒ ĻČżņ¢æņØś ļ│æĒā£ņāØļ”¼ļź╝ ņÖäņĀäĒ׳ ļ│ĆĒÖöņŗ£ĒéżļŖö ĻĘĖņĢ╝ļ¦ÉļĪ£ Ēśüļ¬ģņĀüņØĖ ļ░£Ļ▓¼ņØ┤ļØ╝ ņ╣ŁĒĢĀ ņłś ņ׳ļŗż[4-6].
ļ╣äļĪØ ņ£äņĢö Ļ░äņØś ņŚ░Ļ┤ĆņŚÉņä£ļŖö ĒŚ¼ļ”¼ņĮöļ░ĢĒä░ ĒīīņØ╝ļĪ£ļ”¼ Ļ░ÉņŚ╝ņØ┤ ņäĖĻ│äļ│┤Ļ▒┤ĻĖ░ĻĄ¼ņŚÉņä£ ņĀ£1ĻĖē ļ░£ņĢöņØĖņ×ÉļĪ£ ņĀĢņØśļÉśĻĖ░ļŖö ĒĢśņśĆņ¦Ćļ¦ī, ļ░śļīĆļĪ£ ĒŚ¼ļ”¼ņĮöļ░ĢĒä░ļŖö ĒĢśļéśņØś Ļ│ĄņāØņ▓┤ņØ╝ ņłś ņ׳ļŗż. ļśÉ ņÖäņĀäĒĢ£ ņĀ£ĻĘĀņØĆ ņé¼ņŗżņāü ļČłĻ░ĆļŖźĒĢśļ®░, ļ¦ÄņØĆ ņ×äņāüņŚÉņä£ ļ¬©ļōĀ ĻĘĀņØä Ļ╝Ł ņĀ£ĻĘĀņØä ņŗ£ņ╝£ņĢ╝ ĒĢ£ļŗżļŖö ļ¬ģĒÖĢĒĢ£ Ļ▓░ļĪĀņØ┤ ņĢäņ¦üĻ╣īņ¦ĆļŖö ļé┤ļĀżņ¦Ćņ¦Ć ņĢŖņĢä ņ¦ĆĻĖłļÅä ņØ┤ļź╝ ĻĘ£ļ¬ģĒĢśĻĖ░ ņ£äĒĢ£ ņłśļ¦ÄņØĆ ņŚ░ĻĄ¼Ļ░Ć ņ¦äĒ¢ēļÉśĻ│Ā ņ׳ļŗż. ĒĢśņ¦Ćļ¦ī ņ£ä, ņŗŁņØ┤ņ¦Ćņן ĻČżņ¢æĻ│╝ Ļ░ÖņØĆ ņ¢æņä▒ ņ¦łĒÖśņŚÉņä£ļŖö ĒŚ¼ļ”¼ņĮöļ░ĢĒä░ ĒīīņØ╝ļĪ£ļ”¼ņØś ĒÖ£ļÅÖņØä ņĀ£ĒĢ£ĒĢśĻ▒░ļéś Ļ░ÉņŚ╝ņŚÉ ļö░ļźĖ ņĀ£ļ░ś ļ│æĒā£ņāØļ”¼ļź╝ ņĪ░ņĀłĒĢśĻ│Ā, ņŚ╝ņ”Ø ĒśĖņĀäņØä ņ£äĒĢ£ ņĀ£ĻĘĀ ņ╣śļŻīļź╝ ĒĢśļŖö Ļ▓āņØ┤ ņŚ¼ļŖÉ ļŗżļźĖ ņ£ä ņĀÉļ¦ē ņ×ÉĻĘ╣ņØĖņ×ÉņŚÉ ļīĆĒĢ£ Ļ▓āļ│┤ļŗż ņÜ░ņäĀļÉśļŖö ņ╣śļŻī ļ¬®Ēæ£ļĪ£ ĒÖĢļ”ĮļÉśņŚłļŗż[7,8]. ļ╣äļĪØ ņ£äņĢöĻ│╝ņØś ņŚ░Ļ┤ĆņŚÉ ļīĆĒĢ┤ņä£ļŖö ņĢäņ¦üĻ╣īņ¦ĆļŖö ņĢĮĻ░ä ņāüļ░śļÉ£ ļ░░Ļ▓ĮņŚÉļÅä ļČłĻĄ¼ĒĢśĻ│Ā, ņ£ä ņ¦łĒÖśņØä ņ£Āļ░£ņŗ£Ēé¼ ņłś ņ׳ļŖö ņŚ¼ļ¤¼ Ļ░Ćņ¦Ć Ļ│ĄĻ▓®ņØĖņ×É, ņ”ē ņ£äņé░, ĒØĪņŚ░, ņĢīņĮöņś¼, ņŖżĒŖĖļĀłņŖż, ļŗ┤ņ”Öņé░, Ļ░üņóģ ņĢĮļ¼╝ ļō▒ ļŗżņ¢æĒĢ£ ņÜöņØĖ ņżæņŚÉņä£ļŖö ĒŚ¼ļ”¼ņĮöļ░ĢĒä░ ĒīīņØ╝ļĪ£ļ”¼ ņĀ£ņ¢┤ ļśÉļŖö ņĀ£ĻĘĀņØ┤ ņŚ¼ņĀäĒ׳ Ēü░ ļ╣äņżæņØä Ļ░Ćņ¦ĆĻ│Ā ņ׳ļŗż[9]. ņØĖĻ░äņØś Ļ▓ĮņÜ░ ņĢĮĻ░ä ļ¬ģĒÖĢĒĢśņ¦ĆļŖö ņĢŖņ¦Ćļ¦ī ĒŚ¼ļ”¼ņĮöļ░ĢĒä░ ĻĘĀņŻ╝ņŚÉ ļīĆĒĢ£ ņĪ░ņĀł Ļ░ĆņØ┤ļō£ļØ╝ņØĖņØ┤ ņ׳ļŖö Ļ▓āņŚÉ ļ╣äĒĢśņŚ¼ Ļ░ĆņČĢņØ┤ļéś ļÅÖļ¼╝ņŚÉņä£ļŖö ņØ┤ļ¤░ Ļ░ĆņØ┤ļō£ļØ╝ņØĖņØ┤ ņŚåĻ│Ā, ņĢäņ¦üļÅä ņāüļŗ╣ĒĢ£ ņłśņżĆņØś ĒÅÉņé¼Ļ░Ć ļ¼ĖņĀ£Ļ░Ć ļÉśĻ│Ā ņ׳ļŖö ļÅ╝ņ¦ĆņØś ņśłņÖĆ Ļ░ÖņØ┤ ĒŚ¼ļ”¼ņĮöļ░ĢĒä░ ĒīīņØ╝ļĪ£ļ”¼ Ļ░ÉņŚ╝ņØ┤ ņŗ¼Ļ░üĒĢśĻ▓ī Ļ▒┤Ļ░ĢņØä ĒĢ┤ņ╣śļŖö ņÜöņØĖņØ┤ļØ╝ ĒĢĀ ņłś ņ׳ļŗż. ĻĘĖļ¤¼ļ»ĆļĪ£ ņé¼ļ×ī ņØ┤ņāüņ£╝ļĪ£ ņČĢņé░ ļÅÖļ¼╝ņŚÉĻ▓ī ĒŚ¼ļ”¼ņĮöļ░ĢĒä░ ĒīīņØ╝ļĪ£ļ”¼ Ļ░ÉņŚ╝ņØĆ ļśÉ ĒĢśļéśņØś ĒĢ┤Ļ▓░ļÉśņ¢┤ņĢ╝ ĒĢśļŖö ņŗ£ĻĖēĒĢ£ ļ»ĖņČ®ņĪ▒ ņØśļŻī ņłśņÜö(unmet medical needs)ņØĖ Ļ▓āņØ┤ļŗż[10,11].
ĻĘĖļ¤░ļŹ░ ņØ┤ļ¤¼ĒĢ£ ņ”ØĻ░ĆļÉ£ Ļ│ĄĻ▓®ņØĖņ×ÉņÖĆ ĒĢ©Ļ╗ś ĒŚ¼ļ”¼ņĮöļ░ĢĒä░ ĒīīņØ╝ļĪ£ļ”¼ Ļ░ÉņŚ╝ ņŗ£ņŚÉļŖö ļ░®ņ¢┤ņØĖņ×ÉņØś ņĢĮĒÖöĻ░Ć ĻČżņ¢æņØś ļ░£ņāØ ļ░Å ņĢöĒÖö ļō▒ņŚÉļÅä Ļ┤ĆņŚ¼ĒĢśļŖöļŹ░ ņŗż ņśłļź╝ ļōżļ®┤, ņ£äņĀÉņĢĪņØś ņĢĮĒÖöļéś ņ£äņĀÉļ¦ē ņäĖĒż ņ×¼ņāØņØś ņ¦ĆņŚ░ ĻĘĖļ”¼Ļ│Ā ļČłņČ®ļČäĒĢ£ ņäĖĒż ļ│┤ĒśĖ ņ×æņÜ® ļō▒ļÅä ņ£ä ņ¦łĒÖś ļ░£ņāØņŚÉ ņżæņÜöĒĢśĻĖ░ ļĢīļ¼ĖņŚÉ ņ×äņāüņŚÉņä£ļŖö ĒŚ¼ļ”¼ņĮöļ░ĢĒä░ ĒīīņØ╝ļĪ£ļ”¼ņØś ņĪ░ņĀłĻ│╝ ĒĢ©Ļ╗ś ņØ┤ļōż ļ░®ņ¢┤ņØĖņ×É ņ”ØĻ░ĢņØ┤ ņ╣śļŻīņØś ļśÉ ļŗżļźĖ ņČĢņØä ņ░©ņ¦ĆĒĢ£ļŗż[12,13]. ņ”ē, ņ£äņĀÉņĢĪņØś ņ£Āņ¦Ć, ņ£äņĀÉļ¦ē ņāüĒö╝ņäĖĒżņØś ņ×¼ņāØ ļ░Å ņ£Āņ¦Ć, ņ£äĒśłļźśļ¤ē ņ£Āņ¦Ć ļ░Å ņ£ä ļ│┤ĒśĖņŗĀĻ▓Į Ļ░ĢĒÖö ļ░Å prostaglandin (PG) ņ£Āņ¦Ć ļō▒ņØ┤ ņĢäņŻ╝ ņżæņÜöĒĢ£ ļ░®ņ¢┤ņØĖņ×É ņ▓┤Ļ│äņØĖļŹ░, ņØ┤ ņżæņŚÉņä£ ĒŖ╣Ē׳ ņ£äņĀÉņĢĪ ņāØņä▒ ļ░Å ņĀÉņĢĪ ņåŹ PG ņ£Āņ¦ĆĻ░Ć ņĢäņŻ╝ ĻĘ╝ļ│ĖņĀüņØ┤ļ®░, ļŹö ĻĄ¼ņ▓┤ņĀüņ£╝ļĪ£ļŖö PGļź╝ ĒżĒĢ©ĒĢ£ ņ£äņĀÉņĢĪ ņāØņä▒ ņ”ØĻ░ĆĻ░Ć ņżæņÜöĒĢśļŗżĻ│Ā ĒĢśĻ▓Āļŗż[14].
ņĀĆņ×ÉļōżņØĆ ņØ┤ļ»Ė ĻĄ¼ņåŹ ļ░Å ņ╣©ņłś ņŖżĒŖĖļĀłņŖż(water immersion restraint stress)ņŚÉ ļģĖņČ£ņŗ£ņ╝£ ņ£äĻČżņ¢æņØä ņ£Āļ░£ņŗ£Ēé© ļ¬©ļŹĖņŚÉņä£ threonine Ļ│╝ļ”ĮņØ┤ ņĀÉņĢĪ ņāØņä▒ ņ”ØĻ░Ć ļ░Å ĒĢŁņŚ╝ņ”Ø/ĒĢŁņé░ĒÖö ņ×æņÜ® ļō▒ņŚÉ ĻĘ╝Ļ▒░ĒĢśņŚ¼ ņ£ĀņØśĒĢ£ ĒĢŁĻČżņ¢æ ĒÜ©Ļ│╝ļź╝ ļéśĒāĆļé┤ļŖö Ļ▓āņØä ņ”Øļ¬ģĒĢśņśĆļŗż[15]. ņØ┤ņŚÉ ĒŚ¼ļ”¼ņĮöļ░ĢĒä░ ĒīīņØ╝ļĪ£ļ”¼ Ļ░ÉņŚ╝ ņŗ£ threonine Ļ│╝ļ”ĮņØś ļ│┤ĒśĖ ĒÜ©ļŖźņØĆ ļ¼╝ļĪĀ threonineņØä ļ¦īļōżņ¢┤ ļé┤ļŖö Corynebacterium glutamicum (C. glutamicum)ņØä ĒżĒĢ©ĒĢ£ Corynebacterium species (sp.) ĻĘĀņŻ╝ņÖĆņØś ļ│æĒĢ® Ēł¼ņŚ¼ņŚÉ ļö░ļØ╝ ĒĢŁĒŚ¼ļ”¼ņĮöļ░ĢĒä░ ĒīīņØ╝ļĪ£ļ”¼ ĒÜ©ļŖźņØĆ ļ¼╝ļĪĀ Ļ░üņóģ ĒŚ¼ļ”¼ņĮöļ░ĢĒä░ ĒīīņØ╝ļĪ£ļ”¼ ņŚ░Ļ┤Ć ļ│æĒā£ņāØļ”¼ĒĢÖņĀü ļ░śņØæņØ┤ Ļ░ÉņåīļÉśļŖöņ¦Ć ļō▒ņØä ĻĘ£ļ¬ģĒĢśĻ│Ā, ļśÉĒĢ£ threonine Ļ│╝ļ”ĮņØĆ Ēśäņ×¼ ņé¼ļ×īņØä ļīĆņāüņ£╝ļĪ£ ĒĢśļŖö ņŗØņØ┤ ņä▒ļČäļ│┤ļŗżļŖö Ļ░ĆņČĢņØ┤ļéś ļÅÖļ¼╝ļōżņŚÉĻ▓īņä£ ņ£äņן ļ│┤ĒśĖ ĒÜ©ļŖźņØ┤ ņØĖņĀĢļÉśņ¢┤ ņé¼ļŻī ļō▒ņ£╝ļĪ£ ņé¼ņÜ®ļÉśĻ│Ā ņ׳ļŖö ļ░ö[16-18], ļ│Ė ņŚ░ĻĄ¼ļź╝ ĒåĄĒĢśņŚ¼ ĒŚ¼ļ”¼ņĮöļ░ĢĒä░ ĒīīņØ╝ļĪ£ļ”¼ Ļ░ÉņŚ╝ņŚÉ ļīĆĒĢ£ threonine Ļ│╝ļ”ĮņØś ņ£äņן ļ│┤ĒśĖ ĒÜ©ļŖźņŚÉ ļīĆĒĢśņŚ¼ ļŹö ĒÖĢņŗżĒĢ£ ĻĘ╝Ļ▒░ļź╝ ņĀ£Ļ│ĄĒĢśĻĖ░ ņ£äĒĢśņŚ¼ ļ│Ė ņŚ░ĻĄ¼ļź╝ ņŗ£Ē¢ēĒĢśņśĆļŗż.
ļīĆņāü ļ░Å ļ░®ļ▓Ģ
1. ņäĖĒżņŻ╝ ļ░Å ņäĖĒż ļ░░ņ¢æ
ņĀĢņāü wistar rat ņ£äņĀÉļ¦ē ņäĖĒżņŻ╝ņØĖ RGM-1 ņäĖĒżļŖö 10%ņØś fetal bovine serumņØ┤ ĒżĒĢ©ļÉ£ DulbeccoŌĆÖs modified essential medium (DMEM)Ļ│╝ Ham F12 Ēś╝ĒĢ®ļ░░ņ¦ĆņŚÉņä£ 37Ōäā ņäĖĒż ļ░░ņ¢æĻĖ░(95% air, 5% CO2)ļĪ£ ļ░░ņ¢æĒĢśņśĆļŗż. ļ│Ė ņäĖĒżņŻ╝ļŖö ņØ╝ļ│Ė Tsukuba ļīĆĒĢÖņØś Matsui ĻĄÉņłśņŚÉ ņØśĒĢśņŚ¼ ņłśļ”ĮļÉśņŚłļŖöļŹ░ ļÅÖņØś ĒøäņŚÉ ļ│Ė ņŚ░ĻĄ¼ļź╝ ņ£äĒĢśņŚ¼ ņĀ£Ļ│ĄļÉśņŚłļŗż.
2. ĒŚ¼ļ”¼ņĮöļ░ĢĒä░ ĒīīņØ╝ļĪ£ļ”¼ ĻĘĀņŻ╝ ļ░Å ņäĖĒż Ļ░ÉņŚ╝
ņØ┤ ņŗżĒŚśņŚÉ ņé¼ņÜ®ļÉ£ ĒŚ¼ļ”¼ņĮöļ░ĢĒä░ ĒīīņØ╝ļĪ£ļ”¼ ĻĘĀņŻ╝(cytotoxin-associated gene A+ strain, NCTC 11637)ļŖö American Type Culture Collection (ATCC, Rockville, MD, USA)ņ£╝ļĪ£ļČĆĒä░ ĻĄ¼ņ×ģĒĢśņśĆļŗż. ĻĘĀņŻ╝ļŖö 5% bovine calf serumĻ│╝ ĒĢŁņāØņĀ£Ļ░Ć ņ▓©Ļ░ĆļÉ£ brucella brothņŚÉņä£ 1├Ś108 CFU/mL (optical density [OD] 600=1)ņØ┤ ļÉĀ ļĢīĻ╣īņ¦Ć 10% CO2 ņĪ░Ļ▒┤ņŚÉņä£ ņ¦äĒāĢ ļ░░ņ¢æĒĢśņśĆļŗż. ņäĖĒżņŚÉņä£ņØś Ļ░ÉņŚ╝ ļŗżņżæļÅä(multiplicity of infection, MOI)ļŖö 100:1ļĪ£ ņé¼ņÜ®ĒĢśņśĆļŗż. ĒĢŁĒŚ¼ļ”¼ņĮöļ░ĢĒä░ ĒīīņØ╝ļĪ£ļ”¼ļŖź ņĖĪņĀĢņŚÉļŖö disk diffusion assayņÖĆ ĻĘĀņŻ╝ ļ░░ņ¢æņĢĪņØś OD ņĖĪņĀĢņØä ĒåĄĒĢ£ ņä▒ņן ņāüĒā£ļź╝ ņĖĪņĀĢĒĢśļŖö ļæÉ Ļ░Ćņ¦Ć ļ░®ļ▓ĢņØä ļ¬©ļæÉ ņé¼ņÜ®ĒĢśņśĆļŗż. ļ│Ė ņŚ░ĻĄ¼ļź╝ Ļ░£ņŗ£ĒĢśĻĖ░ ņĀä ļ░░Ļ▓Į ņŚ░ĻĄ¼ļĪ£ ņŗ£Ē¢ēĒĢ£ Supplementary Fig. 1ņŚÉņä£ņÖĆ Ļ░ÖņØ┤ disk ņŻ╝ņ£äņØś ĒĢŁĒŚ¼ļ”¼ņĮöļ░ĢĒä░ ĒīīņØ╝ļĪ£ļ”¼ļŖźņŚÉ ņØśĒĢ£ ņÜ®Ēśł(hemolysis) Ļ▓░Ļ│╝ņŚÉ ļö░ļØ╝ ņāØņä▒ Ļ┤Ćņ░░ļÉśļŖö Ēł¼ļ¬ģĒĢ£ haloļź╝ ņĖĪņĀĢĒĢśņśĆļŗż. ThreonineņØś ļåŹļÅäņŚÉ ļö░ļØ╝ ņĢäņŻ╝ ļŗżņ¢æĒĢ£ Ēü¼ĻĖ░ņØś ĒĢŁĒŚ¼ļ”¼ņĮöļ░ĢĒä░ ĒīīņØ╝ļĪ£ļ”¼ļŖźņŚÉ ņØśĒĢ£ Ēł¼ļ¬ģĒĢ£ haloĻ░Ć ĒśĢņä▒ļÉśņŚłĻ│Ā, ņØ┤ ņŗżĒŚśņØä 3ļ▓ł ņØ┤ņāü ļ░śļ│ĄĒĢśņŚ¼ ĒÅēĻĘĀņĀü ĒĢŁĒŚ¼ļ”¼ņĮöļ░ĢĒä░ ĒÜ©ļŖźņØä ĻĘ£ņĀĢĒĢśņśĆļŗż. ņĀĆņ×ÉļōżņØĆ ņśłļ╣ä ņŚ░ĻĄ¼ļź╝ ĒåĄĒĢśņŚ¼ ļŗżņ¢æĒĢ£ ļåŹļÅäņØś threonineņØś ĒĢŁĒŚ¼ļ”¼ņĮöļ░ĢĒä░ ĒīīņØ╝ļĪ£ļ”¼ ĒÜ©ļŖźņØä ĒŚ¼ļ”¼ņĮöļ░ĢĒä░ ĒīīņØ╝ļĪ£ļ”¼ ļ░░ņ¢æ ņĀäņÜ® blood agar plateņŚÉņä£ ņĖĪņĀĢĒĢśņŚ¼ ļ│Ė Ļ▓░Ļ│╝ ĒĢŁĒŚ¼ļ”¼ņĮöļ░ĢĒä░ ĒÜ©ļŖźņØä 60%, 65% ļ░Å 70% ļåŹļÅäņŚÉņä£ Ļ┤Ćņ░░ņØä ĒĢśņśĆĻĖ░ņŚÉ, ļ│Ė ņŚ░ĻĄ¼ņŚÉņä£ļŖö threonineņØä ņāØņä▒ĒĢśļŖö C. glutamicum ĻĘĀņŻ╝ņÖĆņØś ļ│æĒĢ® Ēł¼ņŚ¼ņŚÉ ļö░ļźĖ ņĀ£ļ░ś ĒŚ¼ļ”¼ņĮöļ░ĢĒä░ ņĀ£ĒĢ£ ĻĖ░ļŖźņØä ņŚ░ĻĄ¼ĒĢ£ Ļ▓āņØ┤ļŗż. ļ│Ė ņŚ░ĻĄ¼ņŚÉņä£ļŖö ņØ┤ļ¤¼ĒĢ£ ļ░░Ļ▓Į ņŚ░ĻĄ¼ņŚÉ ĻĖ░ļ░śĒĢśņŚ¼ Table 1ņŚÉ ņĀ£ņŗ£ĒĢ£ Ļ▓āĻ│╝ Ļ░ÖņØ┤ ņ┤Ø 8Ļ░£ņØś ĻĄ░ņ£╝ļĪ£ ĻĄ¼ļČäĒĢśņŚ¼ threonine ļŗ©ļÅģĻ│╝ ņŚ¼ļ¤¼ Ļ░Ćņ¦Ć Corynebacterium sp. ņÖĆņØś ļ│æĒĢ®ņ£╝ļĪ£ ĻĄ░ņØä ĻĄ¼ņä▒ĒĢśņśĆļŗż.
3. MTT assayļź╝ ĒåĄĒĢ£ ņäĖĒż ņāØņĪ┤ ņĖĪņĀĢ
ņäĖĒżļź╝ 48-well dishņŚÉ 1├Ś104Ļ░£/wellļĪ£ ļÅäĒżĒĢ£ ļŗżņØī DMEM ļ░░ņ¦ĆņŚÉņä£ ļ░░ņ¢æĒĢśņśĆļŗż. ņØ┤Ēøä Ļ░üĻ░üņØś ņŗ£ņĢĮņØä ņØ╝ņĀĢ ņŗ£Ļ░ä ņ▓śļ”¼ĒĢśņŚ¼ MTT solution (ņĄ£ņóģ ļåŹļÅä 1 mg/mL)ņØä Ļ░ĆĒĢśĻ│Ā 2ņŗ£Ļ░ä ļÅÖņĢł ļ░śņØæĒĢ£ Ēøä dimethyl sulfoxideļĪ£ ņÜ®ĒĢ┤ņŗ£Ēé© Ēøä 540 nmņŚÉņä£ ODļź╝ microplate readerļĪ£ ņĖĪņĀĢĒĢśņśĆļŗż.
4. Reverse transcription-PCR ļČäņäØ
RNAļŖö TRIzol (Gibco BRL, Rockville, MD, USA)ņØä ņØ┤ņÜ®ĒĢśņŚ¼ ņČöņČ£ĒĢśņśĆņ£╝ļ®░, ņČöņČ£ļÉ£ RNAļŖö Moloney murine leukemia virus ņŚŁņĀäņé¼ĒÜ©ņåī(Perkin Elmer, Morrisville, NC, USA)ļź╝ ņØ┤ņÜ®ĒĢśņŚ¼ cDNAļĪ£ ĒĢ®ņä▒ĒĢśņśĆļŗż. PCR ļČäņäØņŚÉ ņé¼ņÜ®ļÉ£ Ļ░üĻ░üņØś primer ņä£ņŚ┤ņØĆ Table 1ņŚÉ ļéśĒāĆļé┤ņŚłņ£╝ļ®░, internal standardļĪ£ņä£ glyceraldehyde 3-phosphate dehydrogenase (GAPDH)ļź╝ ņé¼ņÜ®ĒĢśņśĆļŗż.
5. Western blot ļČäņäØ
ļ░░ņ¢æļÉ£ ņäĖĒżļź╝ phosphate buffered saline ņÜ®ņĢĪņ£╝ļĪ£ ņäĖņ▓ÖĒĢ£ Ēøä, cell lysis buffer (150 mM NaCl, 0.5% triton├Ś100, 50 mM tris-HCl, pH 7.4, 25 mM NaF, 20 mM ethyleneglycol-bis [╬▓-aminoethylether]-N, NŌĆÖ-tetraacetic acid, 1 mM dithiothreitol, 1 mM Na3VO4, Protease Inhibitor Cocktail tablet [Boehringer, Manneim, Germany])ļĪ£ ņÜ®ĒĢ┤ņŗ£ņ╝£ protein lysateļź╝ ļ¦īļōżņŚłļŗż. ņØ┤Ļ▓āņØä sodium dodecyl sulfate-polyacrylamide gel electrophoresisļĪ£ ņĀäĻĖ░ ņśüļÅÖĒĢ£ Ēøä ņĀäĻ░£ņŗ£Ēé© ļŗ©ļ░▒ņ¦łņØä PVDF membrane (Gelman Sciences, Ann Arbor, MI, USA)ņŚÉ ņś«Ļ▓© Ļ░üĻ░üņØś primary antibodyņÖĆ secondary antibodyļĪ£ ļ░śņØæņŗ£Ēé© Ēøä, chemoluminescence systemņØä ņØ┤ņÜ®ĒĢśņŚ¼ ļČäņäØĒĢśņśĆļŗż.
6. 2',7'-dichlorofluorescein-diacetate (DCF-DA) for oxidative stress
24 well plateņŚÉ 5├Ś105/wellņØś RGM-1 ņäĖĒżļź╝ ļČäņŻ╝ĒĢśĻ│Ā 37Ōäā ņäĖĒż ļ░░ņ¢æĻĖ░(95% air, 5% CO2)ņŚÉņä£ ļ░żņāł ļ░░ņ¢æ Ēøä Ļ░üĻ░üņØś ņŗ£ņĢĮņØä ņØ╝ņĀĢ ņŗ£Ļ░ä ņ▓śļ”¼ĒĢśņśĆļŗż. DMEMņ£╝ļĪ£ ņäĖĒżļź╝ ņö╗ņ¢┤ļéĖ Ēøä 10 ╬╝g/mL DCF-DA (Sigma-Aldrich Co., St. Louis, MO, USA)ļź╝ ļ░░ņ¦ĆņŚÉ ņ▓©Ļ░ĆĒĢśņŚ¼ ņØĖĒüÉļ▓ĀņØ┤Ēä░ņŚÉņä£ 30ļČä ļÅÖņĢł ļ░░ņ¢æĒĢśņśĆļŗż. ļ░░ņ¢æ Ēøä 4ŌäāņØś phosphate-buffered saline (PBS)ļĪ£ ņö╗ņ¢┤ļéĖ ņäĖĒżļź╝ ĒśĢĻ┤æ Ēśäļ»ĖĻ▓Įņ£╝ļĪ£ Ļ┤Ćņ░░ ļ░Å ņ┤¼ņśüĒĢśņśĆļŗż.
7. TUNEL staining
DNA ļČäņĀł ĒśäņāüņØä TUNELņØś ļ░®ļ▓Ģņ£╝ļĪ£ ņĪ░ņé¼ĒĢśĻĖ░ ņ£äĒĢśņŚ¼ ņäĖĒżļź╝ Lab-Tek chamber slideņŚÉ 1├Ś105Ļ░£/chamberļĪ£ ļČäņŻ╝ĒĢ£ ļŗżņØī 37Ōäā ņäĖĒż ļ░░ņ¢æĻĖ░(95% air, 5% CO2)ļĪ£ ļ░░ņ¢æĒĢśņśĆļŗż. ņØ┤Ēøä Ļ░üĻ░üņØś ņŗ£ņĢĮņØä ņØ╝ņĀĢ ņŗ£Ļ░ä ņ▓śļ”¼ĒĢ£ Ēøä apoptosis detection kit(invitrogen, Denver, CO, USA)ļź╝ ņé¼ņÜ®ĒĢśņŚ¼ ņĀ£ņĪ░ĒÜīņé¼ņØś ņŗżĒŚś ļ░®ļ▓ĢļīĆļĪ£ ņŗ£Ē¢ēĒĢśņśĆļŗż. ļ░░ņ¢æņĢĪņØä ņĀ£Ļ▒░ĒĢśĻ│Ā PBSļĪ£ 3ĒÜī ņäĖņ▓Ö Ēøä 4% paraformaldehydeļź╝ ņ▓©Ļ░ĆĒĢśņŚ¼ ņāüņś©ņŚÉņä£ 20ļČäĻ░ä Ļ│ĀņĀĢĒĢśņśĆļŗż. PBSļĪ£ ļŗżņŗ£ 2ĒÜī ņäĖņ▓Ö Ēøä 0.1% triton X-100ņØä 4ŌäāņŚÉņä£ 5ļČäĻ░ä ņ▓śņ╣śĒĢśĻ│Ā terminal deoxynucleotidyl transferaseņÖĆ nucleotide mixtureļź╝ Ēś╝ĒĢ®ĒĢśņŚ¼ ļ¦īļōĀ ņÜ®ņĢĪņ£╝ļĪ£ 37ŌäāņŚÉņä£ ļ╣øņØä ņ░©ļŗ©ĒĢ£ Ēøä 60ļČäĻ░ä ļ░śņØæņŗ£ņ╝░ļŗż. PBSļĪ£ ņäĖņ▓ÖĒĢśĻ│Ā ļé£ Ēøä, ĒśĢĻ┤æ Ēśäļ»ĖĻ▓Įņ£╝ļĪ£ ņäĖĒżņ×Éļ®Ėņé¼ļź╝ Ļ┤Ćņ░░ĒĢśņśĆļŗż.
Ļ▓░ Ļ│╝
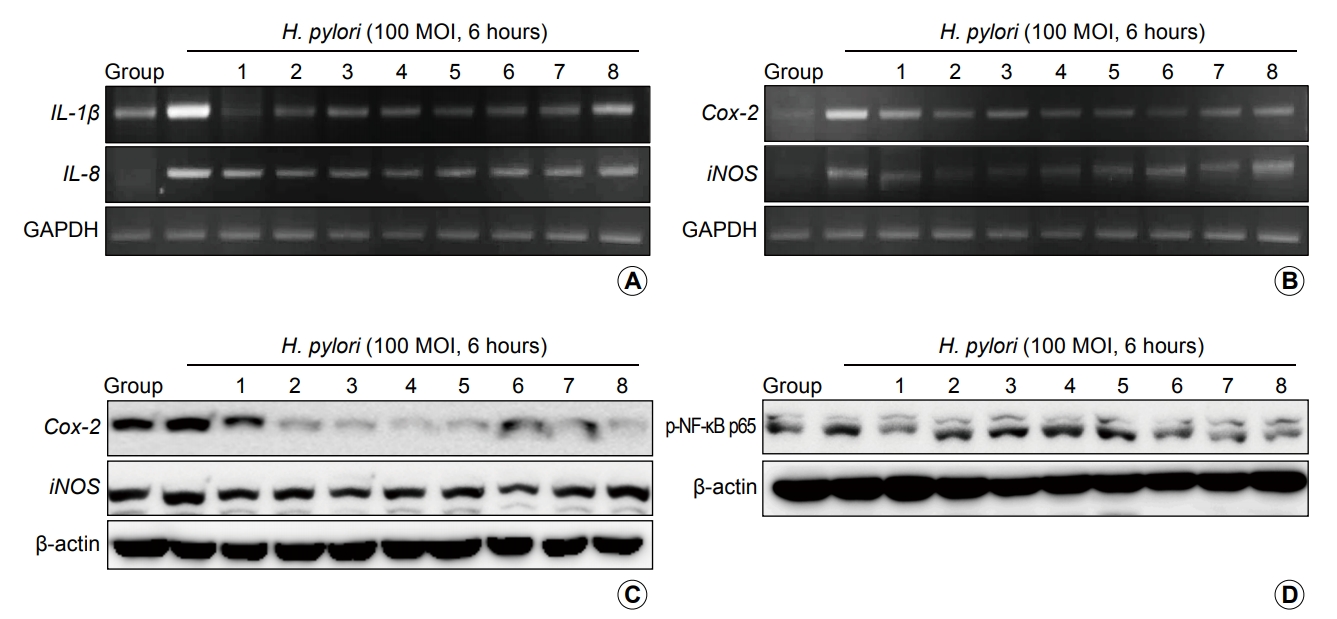
1. Ļ│╝ļ”Į threonineĻ│╝ C. glutamicum ļ│æĒĢ®ņŚÉ ņØśĒĢ£ ĒŚ¼ļ”¼ņĮöļ░ĢĒä░ ĒīīņØ╝ļĪ£ļ”¼ ņŚ░Ļ┤Ć Ļ░üņóģ ņŚ╝ņ”Ø ļ¦żĻ░£ņØĖņ×ÉņØś Ļ░ÉņåīņÖĆ ņØ┤ļź╝ ņĀäņé¼ĒĢśļŖö NF-╬║B ņĀäņé¼ņØĖņ×É ļ╣äĒÖ£ņä▒ĒÖö
Fig. 1AņÖĆ 1BņŚÉņä£ņÖĆ Ļ░ÖņØ┤ 6ņŗ£Ļ░ä ļÅÖņĢł 100 MOI ĒŚ¼ļ”¼ņĮöļ░ĢĒä░ ĒīīņØ╝ļĪ£ļ”¼ļź╝ ņ£äņĀÉļ¦ē ņāüĒö╝ņäĖĒżņŚÉ Ļ░ÉņŚ╝ņŗ£Ēé© Ļ▓ĮņÜ░ ņŚ╝ņ”Ø ļ¦żĻ░£ņØĖņ×ÉņØĖ IL-1╬▓, IL-8, Cox-2, iNOS mRNAĻ░Ć ņ£ĀņØśĒĢśĻ▓ī ņ”ØĻ░ĆļÉ©ņØä Ļ┤Ćņ░░ĒĢĀ ņłś ņ׳ņŚłļŗż(P<0.01). Western blotņ£╝ļĪ£ ļŗ©ļ░▒ņ¦łņŚÉņä£ļÅä ņ£ĀņØśĒĢ£ ņ”ØĻ░Ćļź╝ Ļ┤Ćņ░░ĒĢĀ ņłś ņ׳ņŚłņ£╝ļ®░, ņØ┤ļ¤¼ĒĢ£ ļ│ĆĒÖöļŖö ņØ┤ļź╝ ņżæĻ░£ĒĢśļŖö NF-╬║B p65ņÖĆ Ļ░ÖņØĆ ņĀäņé¼ņØĖņ×ÉņØś ĒÖ£ņä▒ĒÖö ņ”ØĻ░ĆņÖĆ ņŚ░Ļ┤Ćņä▒ņØ┤ ņ׳ņŚłļŗż(P<0.05; Fig. 1). ĻĘĖļ¤¼ļéś ņØ┤ļ¤¼ĒĢ£ ļ░░Ļ▓Į ĒĢśņŚÉ threonineņØ┤ļéś threonineņØä ņāØņä▒ĒĢĀ ņłś ņ׳ļŖö Corynebacteria sp.ņÖĆņØś ņĖĪņĀĢņØä ĒåĄĒĢśņŚ¼ Ļ┤Ćņ░░ĒĢ£ Ļ░üĻ░ü ĻĘĖļŻ╣ Ļ░äņØś ļ│ĆĒÖöļŖö Fig. 1ņŚÉņä£ ļ│┤ņŚ¼ņŻ╝ļō»ņØ┤ ļīĆņĪ░ĻĄ░ņŚÉ ļ╣äĒĢśņŚ¼ threonine Ēł¼ņŚ¼ĻĄ░ņŚÉņä£ ļ¬©ļæÉ ņ£ĀņØśĒĢ£ ļ│ĆĒÖöļź╝ ļ│┤ņŚ¼, Cox-2, iNOSļŖö ļ¼╝ļĪĀ NF-╬║B ļ╣äĒÖ£ņä▒ĒÖöļĪ£ ņØ┤ņ¢┤ņĪīĻ│Ā(P<0.05), ĒŖ╣Ē׳ group 6Ļ│╝ 7ņŚÉņä£ Ļ░Ćņן ņ£ĀņØśĒĢ£ ļ│ĆĒÖöļź╝ ļ│┤ņŚ¼ņŻ╝ņ¢┤ ņØ┤ļ¤¼ĒĢ£ Ļ▓░Ļ│╝ļŖö threonine ļŗ©ļÅģņØ┤ļéś ņØ┤ļź╝ ņāØņä▒ĒĢĀ ņłś ņ׳ļŖö CorynebacteriumĻĘĀņŻ╝ņØś brothņÖĆņØś ļ│æĒĢ®ņ£╝ļĪ£ ļŹöņÜ▒ ļæÉļō£ļ¤¼ņ¦ĆļŖö Ļ▓░Ļ│╝ņŚÉ ĻĖ░ņ┤łĒĢśņŚ¼ threonineņØ┤ ņ£äņĀÉļ¦ē ņāüĒö╝ņäĖĒżņŚÉņä£ ĒĢŁĒŚ¼ļ”¼ņĮöļ░ĢĒä░ ĻĖ░ļŖźņØä ņ£ĀļÅäĒĢ©ņØä ņĢī ņłś ņ׳ņŚłļŗż.
2. Ļ│╝ļ”Į threonineĻ│╝ C. glutamicum ļ│æĒĢ®ņŚÉ ņØśĒĢ£ ĒŚ¼ļ”¼ņĮöļ░ĢĒä░ ĒīīņØ╝ļĪ£ļ”¼ ņŚ░Ļ┤Ć oxidative stress ņĢĮĒÖö ĒÜ©ļŖź
Fig. 2AņÖĆ 2BļŖö ĒŚ¼ļ”¼ņĮöļ░ĢĒä░ ĒīīņØ╝ļĪ£ļ”¼ Ļ░ÉņŚ╝ Ēøä oxidative stressņÖĆ ņŚ░Ļ┤ĆļÉ£ HIF-1╬▒ņÖĆ acute phase responseļź╝ ņĀäņé¼ĒĢśļŖö c-FosņÖĆ c-JunņØś ļ│ĆĒÖöļź╝ Western blotņ£╝ļĪ£ ņĖĪņĀĢĒĢ£ Ļ▓āņ£╝ļĪ£, ĒŚ¼ ļ”¼ņĮöļ░ĢĒä░ ĒīīņØ╝ļĪ£ļ”¼ Ļ░ÉņŚ╝(6ņŗ£Ļ░ä, 100 MOI)ņŚÉ ņØśĒĢśņŚ¼ ņ£ĀņØśĒĢśĻ▓ī acute phase response geneņØ┤ ņ”ØĻ░ĆĒĢśļ®░, ļÅÖņŗ£ņŚÉ HIF-1╬▒ņØś ņ”ØĻ░Ćļź╝ Ļ┤Ćņ░░ĒĢĀ ņłś ņ׳ņŚłļŗż. ļ░śļ®┤ņŚÉ threonine ļŗ©ļÅģņØ┤ļéś ņāØņä▒ ĻĘĀņŻ╝ņÖĆņØś ļ│æĒĢ® Ēł¼ņŚ¼ņŚÉ ļö░ļØ╝ ņ£ĀņØśĒĢ£ ļ│ĆĒÖöļź╝ ļ│┤ņŚ¼ņŻ╝ņŚłļŖöļŹ░, ĻĄ¼ņ▓┤ņĀüņ£╝ļĪ£ ĒŖ╣Ē׳ group 6Ļ│╝ group 7ņŚÉņä£ ņĢäņŻ╝ ņ£ĀņØśĒĢ£ HIF-1╬▒ņØś Ļ░Éņåīļź╝ ļ│┤ņØĖ ļ░śļ®┤ņŚÉ c-JunĻ│╝ c-FosļŖö group 3ņŚÉņä£ ņ£ĀņØśĒĢ£ Ļ░Éņåīļź╝, c-FosņØś Ļ▓ĮņÜ░ņŚÉļŖö ļ░śļīĆļĪ£ group 6ņŚÉņä£ ņ£ĀņØśĒĢ£ ņ”ØĻ░Ćļź╝ ļ│┤ņŚ¼ņä£ ņØ┤ļ¤¼ĒĢ£ acute phase responderņØś ĒĢĄļé┤ ņØ┤ļÅÖņØś ļ│ĆĒÖöļŖö threonine Ēł¼ņŚ¼Ļ░Ć ĒŚ¼ļ”¼ņĮöļ░ĢĒä░ ĒīīņØ╝ļĪ£ļ”¼ Ļ░ÉņŚ╝ņŚÉ ņ”ēĻ░üņĀüņØĖ ņāØņ▓┤ ļ░śņØæņØä ļ│┤ņŚ¼ņŻ╝ļŖö ņŗØņØ┤ņØĖņ×ÉņØ╝ ņłś ņ׳ņØīņØä ņŗ£ņé¼ĒĢśļŖö ņåīĻ▓¼ņØ┤ļØ╝ ĒĢśĻ▓Āļŗż. ņØ┤ļ¤¼ĒĢ£ ņāØņ▓┤ ļ░śņØæ, ĒŖ╣Ē׳ oxidative stressļź╝ ņŗ£Ļ░üĒÖöĒĢśĻĖ░ ņ£äĒĢśņŚ¼ DCF-DA probeļź╝ ĒÖ£ņÜ®ĒĢ£ DCF ļ░£Ēśä ļ░śņØæņØä confocal imagerļĪ£ ļČäņäØĒĢ£ Fig. 2CņØś Ļ▓░Ļ│╝ļź╝ ļ│┤ļ®┤ ĒŚ¼ļ”¼ņĮöļ░ĢĒä░ ĒīīņØ╝ļĪ£ļ”¼ Ļ░ÉņŚ╝ Ēøä ņ£ĀņØśĒĢśĻ▓ī ņ”ØĻ░ĆļÉ£ DCFļŖö group 1, 3Ļ│╝ 6ņŚÉņä£ ĒåĄĻ│äņĀüņ£╝ļĪ£ ņ£ĀņØśĒĢśĻ▓ī Ļ░ÉņåīļÉ©ņØä Ļ┤Ćņ░░ĒĢĀ ņłś ņ׳ņŚłļŗż(P<0.05). Ļ▓░ļĪĀņĀüņ£╝ļĪ£ threonine ļŗ©ļÅģņØ┤ļéś ņØ┤ļź╝ ņāØņä▒ĒĢśļŖö CorynebacteriumņØś Ēł¼ņŚ¼ļŖö ĒŚ¼ļ”¼ņĮöļ░ĢĒä░ ĒīīņØ╝ļĪ£ļ”¼ņŚÉ ņØśĒĢ£ oxidative stressļéś hypoxia responseļź╝ ņ”ēĻ░üņĀüņ£╝ļĪ£ ļīĆņ▓śĒĢ©ņØä ņĢī ņłś ņ׳ņŚłļŗż.
3. Ļ│╝ļ”Į threonineĻ│╝ C. glutamicum ļ│æĒĢ®ņŚÉ ņØśĒĢ£ ĒŚ¼ļ”¼ņĮöļ░ĢĒä░ ĒīīņØ╝ļĪ£ļ”¼ ņŚ░Ļ┤Ć Nrf-2 ļ¦żĻ░£ ņäĖĒż ļ│┤ĒśĖņ×æņÜ®
Fig. 3AņŚÉņä£ļŖö ļīĆĒæ£ņĀüņØĖ host cytoprotective factorņØĖ ĒĢŁņé░ĒÖö ĻĖ░ļŖźņØä ļ│┤ņØ┤ļŖö heme oxygenase-1 (HO-1)Ļ│╝ heat shock protein 70 (HSP70) mRNAļź╝ ĒŚ¼ļ”¼ņĮöļ░ĢĒä░ ĒīīņØ╝ļĪ£ļ”¼ Ļ░ÉņŚ╝ ĒøäņÖĆ Ļ░ü ĻĄ░ Ļ░äņØś ļ░£Ēśä ļ│ĆĒÖöļź╝ ņé┤ĒÄ┤ļ│Ė ļ░ö, ĒŚ¼ļ”¼ņĮöļ░ĢĒä░ ĒīīņØ╝ļĪ£ļ”¼ Ļ░ÉņŚ╝ Ēøä ļ░£ņāØĒĢśļŖö oxidative stress ļ░Å ņäĖĒż ņé¼ļ®ĖņŚÉļŖö HO-1 ļ░Å HSP70 mRNAņØś ņ£ĀņØśĒĢ£ Ļ░ÉņåīĻ░Ć Ļ┤Ćņ░░ļÉśņŚłņ£╝ļéś(P<0.001), ļ░śļ®┤ņŚÉ Fig. 3AņÖĆ 3BņŚÉņä£ Ļ┤Ćņ░░ļÉśļō»ņØ┤ group 6ņŚÉņä£ ņ£ĀņØśĒĢ£ Nrf2 ņ”ØĻ░ĆņŚÉ ļö░ļźĖ HO-1ņØś ņ”ØĻ░Ćļź╝ ļ│╝ ņłś ņ׳ņŚłļŗż(P<0.001). HSP70ņØś ļ│ĆĒÖöļŖö group 7 ļ░Å group 8ņŚÉņä£ ļæÉļō£ļ¤¼ņ¦ĆĻ▓ī ņ”ØĻ░ĆĒĢśņśĆļŗż. ĻĘĖļ¤¼ļ»ĆļĪ£ HO-1ņØä ĒżĒĢ©ĒĢ£ ļŗżļźĖ glutathione-s-transferase ļ░Å NADH dehydrogenase quinone-1 (NQO-1)Ļ│╝ Ļ░ÖņØĆ HO-1ņŚÉ ĒżĒĢ©ļÉśļŖö ņĀäļ░śņĀüņØĖ host defense anti-oxidative responseņŚÉ ļīĆĒĢ£ ļ│ĆĒÖöļź╝ ņČöĻ░ĆĒĢśņŚ¼ ņĖĪņĀĢĒĢśņŚ¼ ļ│Ė ļ░ö, Fig. 3CņŚÉņä£ņÖĆ Ļ░ÖņØ┤ threonine ļŗ©ļÅģņØ┤ļéś CorynebacteriumĻ│╝ņØś ļ│æĒĢ®ņØĆ Nrf-2 ĻĖ░ļ░śņØś HO-1 ļ░śņØæņŚÉ ņŻ╝ļĪ£ ĻĖ░ļ░śĒĢśļŖö ĒĢŁņé░ĒÖö ņ×æņÜ®ņØä ļ│┤ņØ┤ļ®░, ņØ┤ļŖö ĒĢŁĒŚ¼ļ”¼ņĮöļ░ĢĒä░ ĒīīņØ╝ļĪ£ļ”¼ņŚÉ ņŻ╝ĒÜ©ĒĢśļŖö ļČäņ×ÉņāØļ¼╝ĒĢÖņĀü ļ░śņØæņ×äņØä ĒÖĢņØĖĒĢĀ ņłś ņ׳ņŚłļŗż.
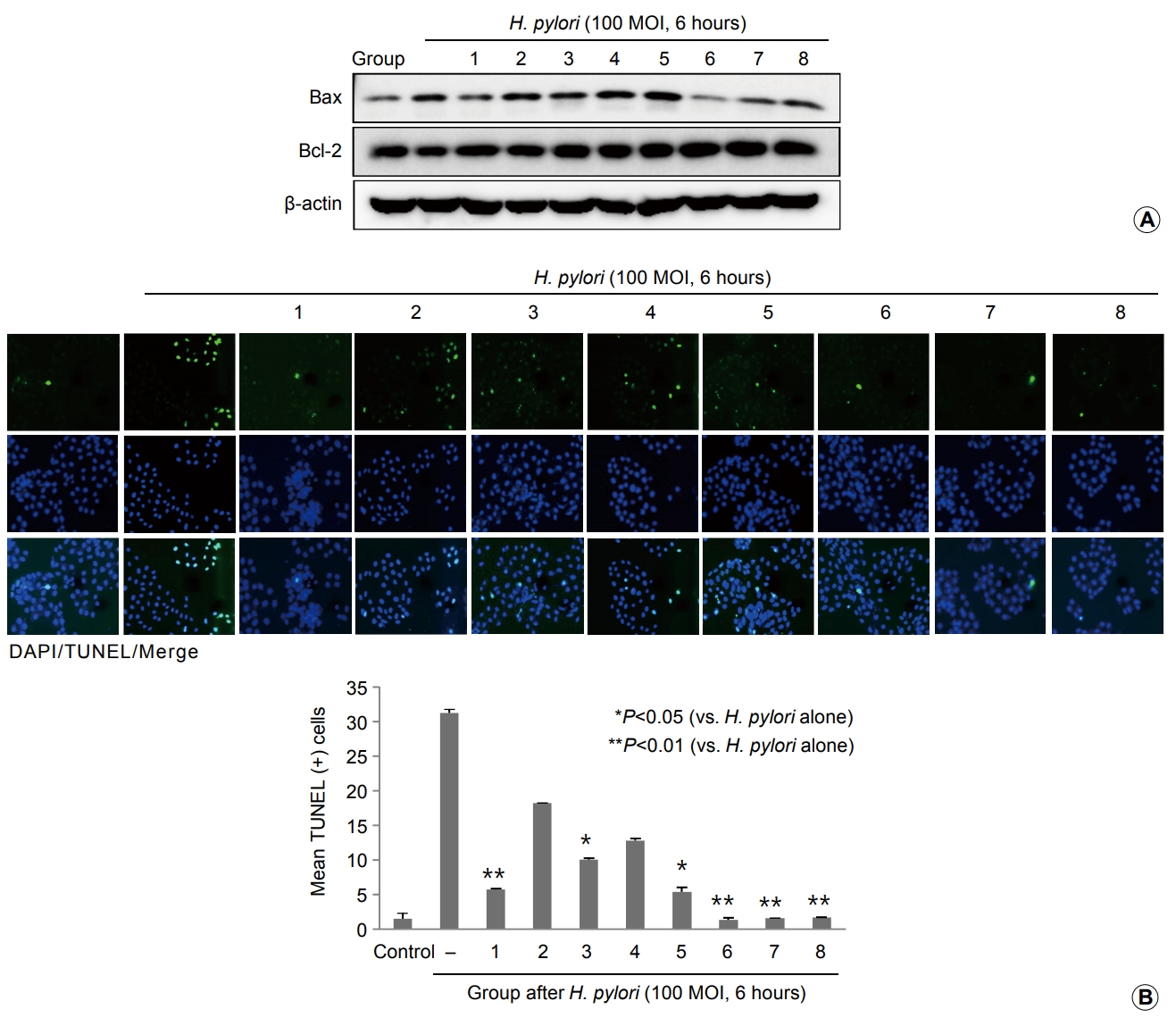
4. Ļ│╝ļ”Į threonineĻ│╝ C. glutamicum ļ│æĒĢ®ņŚÉ ņØśĒĢ£ ĒŚ¼ļ”¼ņĮöļ░ĢĒä░ ĒīīņØ╝ļĪ£ļ”¼ ņŚ░Ļ┤Ć ņäĖĒż ņé¼ļ®Ė ņĀĆĒĢ┤ ĒÜ©ļŖź
ĒŚ¼ļ”¼ņĮöļ░ĢĒä░ ĒīīņØ╝ļĪ£ļ”¼ Ļ░ÉņŚ╝ņØĆ ņäĖĒż ņé¼ļ®ĖņØś ņ£ĀņØśĒĢ£ ņ”ØĻ░ĆņŚÉ ņØśĒĢśņŚ¼ ņĀÉļ¦ē ņåÉņāü ļ░Å ĻČżņ¢æņØä ņ£Āļ░£ĒĢśļŖö Ļ▓āņ£╝ļĪ£ ņל ņĢīļĀżņĀĖ ņ׳ļŗż. Fig. 4AņŚÉņä£ļÅä ĒŚ¼ļ”¼ņĮöļ░ĢĒä░ ĒīīņØ╝ļĪ£ļ”¼ Ļ░ÉņŚ╝ ĒøäņŚÉļŖö ņäĖĒż ņé¼ļ®Ė ņ¢ĄņĀ£ ļŗ©ļ░▒ņØĖ Bcl-2ņØś Ļ░ÉņåīņÖĆ ņäĖĒż ņé¼ļ®Ė ņ£ĀņĀäņ×ÉņØĖ BaxņØś ņ”ØĻ░Ćļź╝ Ļ┤Ćņ░░ĒĢĀ ņłś ņ׳ņŚłĻ│Ā, ņØ┤ļź╝ in situļĪ£ ņ”Øļ╣ÖĒĢśĻĖ░ ņ£äĒĢ£ Fig. 4BņØś TUNEL ņŚ╝ņāēņŚÉņä£ļÅä ĒåĄĻ│äņĀüņ£╝ļĪ£ ņ£ĀņØśĒĢ£ apoptosisņØś ņ”ØĻ░ĆĻ░Ć ĒŚ¼ļ”¼ņĮöļ░ĢĒä░ ĒīīņØ╝ļĪ£ļ”¼ Ļ░ÉņŚ╝ ņØ┤ĒøäņŚÉ Ļ┤Ćņ░░ļÉśņŚłņ£╝ļéś(P<0.01), ļ░śļ®┤ņŚÉ threonine ļŗ©ļÅģņØ┤ļéś Corynebacterium ļÅÖņŗ£ Ēł¼ņŚ¼ĻĄ░ņŚÉņä£ļŖö ĒåĄĻ│äņĀüņ£╝ļĪ£ ņ£ĀņØśĒĢ£ apoptosisņØś Ļ░Éņåīļź╝ Ļ┤Ćņ░░ĒĢĀ ņłś ņ׳ņŚłļŗż(P<0.001; Fig. 5A).
5. Ļ│╝ļ”Į threonineĻ│╝ C. glutamicum ļ│æĒĢ®ņŚÉ ņØśĒĢ£ ĒŚ¼ļ”¼ņĮöļ░ĢĒä░ ĒīīņØ╝ļĪ£ļ”¼ ņŚ░Ļ┤Ć Ļ░üņóģ ņä▒ņןņØĖņ×ÉņØś ņĪ░ņĀłĻĖ░ļŖź
Fig. 5BņŚÉņä£ļŖö ĒŚ¼ļ”¼ņĮöļ░ĢĒä░ ĒīīņØ╝ļĪ£ļ”¼ ņŚ░Ļ┤Ć ņäĖĒż ņä▒ņן, ĒśłĻ┤Ć ņä▒ņן ĻĘĖļ”¼Ļ│Ā Fig. 5AņŚÉņä£ļŖö ņāüĒö╝ņäĖĒż ņ”ØņŗØņØĖņ×ÉņŚÉ ļīĆĒĢśņŚ¼ ņé┤ĒÄ┤ļ│┤ņĢśļŗż. ņØ┤ļ»Ė ļ│┤Ļ│ĀļÉ£ ļ░öņÖĆ Ļ░ÖņØ┤ ĒŚ¼ļ”¼ņĮöļ░ĢĒä░ ĒīīņØ╝ļĪ£ļ”¼ Ļ░ÉņŚ╝ ĒøäņŚÉļŖö EGF, PDGFR, TGF-╬▓ ļ░Å VEGFņØś ņ£ĀņØśĒĢ£ ņ”ØĻ░Ćļź╝ Ļ┤Ćņ░░ĒĢĀ ņłś ņ׳ņŚłļŖöļŹ░(P<0.01), ņØ┤ļ¤¼ĒĢ£ ņåīĻ▓¼ņØĆ ņĢ×ņŚÉņä£ ņØ┤ļ»Ė ņåīĻ░£ļÉ£ ĒŚ¼ļ”¼ņĮöļ░ĢĒä░ ĒīīņØ╝ļĪ£ļ”¼ Ļ░ÉņŚ╝ņŚÉ ļö░ļźĖ ņŚ╝ņ”Ø ņ”ØĻ░ĆņØś Ļ▓░Ļ│╝ļĪ£ ņĀÉļ¦ē ĒŚłĒśł(ischemia) ņ£ĀļÅäņŚÉ ļö░ļźĖ ņØ┤ņ░©ņĀüņØĖ ļ│ĆĒÖöņÖĆ ņØ╝ņ╣śĒĢśļŖö ņåīĻ▓¼ņØ┤ļ®░, ņ”ØĻ░ĆļÉ£ ņØ┤ļōż ņä▒ņןņØĖņ×ÉļŖö ļ¼┤ļČäļ│äĒĢ£ ņĀÉļ¦ē ņ”ØņŗØņØ┤ļéś ņĢöņä▒ ņŚ╝ņ”ØņØä ņĪ░ņןĒĢĀ ņłś ņ׳Ļ│Ā, ņŚŁņ£╝ļĪ£ ņŚ┤ņĢģĒĢ£ ņ£äņĀÉļ¦ē ņŚ╝ņ”Ø ĒÖśĻ▓ĮņØä ļ░śņśüĒĢ┤ņŻ╝ļŖö Ļ▓░Ļ│╝ļØ╝ ĒĢĀ ņłś ņ׳ļŗż. ņØ┤ļ¤¼ĒĢ£ ļ│ĆĒÖöņÖĆ ļŗ¼ļ”¼(P<0.01) threonine ļŗ©ļÅģņØ┤ļéś CorynebacteriumĻ│╝ņØś ļ│æĒĢ® Ēł¼ņŚ¼ļŖö ņØ┤ļź╝ ņ£ĀņØśņä▒ņØ┤ ņ׳Ļ▓ī ņĪ░ņĀłĒĢĀ ņłś ņ׳ņŚłĻ│Ā, ĒŖ╣Ē׳ ņĢ×ņäĀ ņŚ░ĻĄ¼ Ļ▓░Ļ│╝ņÖĆ ņØ╝ņ╣śĒĢśĻ▓ī group 6ņØ┤ļéś group 7ņŚÉņä£ņØś ļŹöņÜ▒ ĒśäņĀĆĒĢ£ ļ│ĆĒÖöļź╝ ļ│┤ņŚ¼, ņĀäņ▓┤ņĀüņ£╝ļĪ£ threonine Ēł¼ņŚ¼ļŖö ĒŚ¼ļ”¼ņĮöļ░ĢĒä░ ĒīīņØ╝ļĪ£ļ”¼ Ļ░ÉņŚ╝ņŚÉ ņØśĒĢ£ ņĀ£ļ░ś ņŚ╝ņ”ØņØ┤ļéś ņĢöĒÖöļź╝ ņÖäĒÖöņŗ£Ēé¼ ņłś ņ׳ļŗżļŖö Ļ▓░Ļ│╝ļź╝ ļ│┤ņŚ¼ņŻ╝ņŚłļŗż. ĻĘĖļ¤¼ļéś ╬▓-catenin ĒÖ£ņä▒ĒÖöņŚÉ ļö░ļźĖ ņäĖĒż ņ”ØņŗØ ņĖĪļ®┤ņŚÉņä£ļŖö ņ£ĀņØśĒĢ£ ļ│ĆĒÖöĻ░Ć Ļ┤Ćņ░░ļÉśņ¦Ć ņĢŖņĢśļŖöļŹ░(Fig. 5B), ņØ┤ļŖö ļ│Ė ņŚ░ĻĄ¼ņØś 100 MOI, 6ņŗ£Ļ░ä Ļ░ÉņŚ╝ņØĆ ņĢäņ¦ü ╬▓-cateninņØś ņ£ĀņØśĒĢ£ ļ│ĆĒÖöļź╝ ņ£ĀļÅäĒĢśļŖö phaseļŖö ņĢäļŗłĻĖ░ ļĢīļ¼ĖņØĖ Ļ▓āņ£╝ļĪ£ ņé¼ļŻīļÉśļéś nuclear fractionņØä ĒÖ£ņÜ®ĒĢ£ ╬▓-cateninņØś nuclear transfer ņŚ░ĻĄ¼ņŚÉņä£ļŖö group 6ņŚÉņä£ ņØ┤ļ»Ė ╬▓-catenin nuclear transferļź╝ ņØ┤ļ»Ė 6ņŗ£Ļ░äņŚÉņä£ļÅä ņĀĆĒĢ┤ņŗ£Ēé¼ ņłś ņ׳ņØīņØ┤ Ļ┤Ćņ░░ļÉśņŚłļŗż.
6. Ļ│╝ļ”Į threonineĻ│╝ Ļ░üņóģ CorynebacteriumņØś ĒĢŁĒŚ¼ļ”¼ņĮöļ░ĢĒä░ ĒÜ©ļŖź
Disc diffusion assay ļ░®ļ▓ĢņØä ņé¼ņÜ®ĒĢśņŚ¼ blood agar plateņŚÉņä£ ĒŚ¼ļ”¼ņĮöļ░ĢĒä░ ĒīīņØ╝ļĪ£ļ”¼ņŚÉ ļīĆĒĢ£ bactericidal ĒÜ©ļŖźņØä ņĖĪņĀĢĒĢśņśĆļŖöļŹ░, ņØ┤ņĀä ņśłļ╣ä ņŗżĒŚś(Supplementary Fig. 1)ņŚÉņä£ threonine ļŗ©ļÅģņ£╝ļĪ£ļÅä ņ£ĀņØśĒĢ£ ĒĢŁĒŚ¼ļ”¼ņĮöļ░ĢĒä░ ĒīīņØ╝ļĪ£ļ”¼ ĒÜ©ļŖźņØ┤ ņ׳ņØīņØä Ļ┤Ćņ░░ĒĢśņśĆļŗż. ņØ┤ļ¤¼ĒĢ£ ļ░░Ļ▓Į ņŚ░ĻĄ¼ļź╝ ĻĘ╝Ļ▒░ļĪ£ ļ│Ė ņŚ░ĻĄ¼ņŚÉņä£ļŖö threonineņØä ņāØņä▒ņŗ£ĒéżļŖö ĻĘĀņŻ╝ņØĖ CorynebacteriaņÖĆņØś ļ│æĒĢ®ņŚÉ ļö░ļźĖ ĒĢŁĒŚ¼ļ”¼ņĮöļ░ĢĒä░ ĒīīņØ╝ļĪ£ļ”¼ ĒÜ©ļŖźņØä ņĖĪņĀĢĒĢśņŚ¼ Fig. 6Ļ│╝ Ļ░ÖņØĆ Ļ▓░Ļ│╝ļź╝ ņ¢╗ņŚłļŗż. Threonine Ļ▓░Ļ│╝ļź╝ ļ░░Ļ▓Įņ£╝ļĪ£ Ēł¼ņŚ¼ņŗ£Ēé© threonineņØĆ CorynebacteriumņŚÉ ņØśĒĢśņŚ¼ ņāØņä▒ņØ┤ ļÉśļ»ĆļĪ£, ļŗżņØīņ£╝ļĪ£ļŖö C. glutamicum, C. ammoniagenes ĻĘĖļ”¼Ļ│Ā ļŗżļźĖ Corynebacterium sp. Ļ░üĻ░üņØś ĻĘĀņŻ╝ ļśÉļŖö ņØ┤ļōż ĻĘĀņŻ╝ļź╝ ļ░░ņ¢æĒĢ£ brothļź╝ ļ│æĒĢ®ĒĢśņŚ¼ Table 2ņÖĆ Ļ░ÖņØĆ ņĪ░ĒĢ®ņ£╝ļĪ£ agar diffusion assay ļ░®ļ▓Ģņ£╝ļĪ£ ĒĢŁĒŚ¼ļ”¼ņĮöļ░ĢĒä░ ĒīīņØ╝ļĪ£ļ”¼ ĒÜ©ļŖźņØä ļ╣äĻĄÉĒĢśņŚ¼ ļ│┤ņĢśļŗż. Fig. 6AņÖĆ Ļ░ÖņØĆ threonine Ļ│╝ļ”ĮņØś ĒĢŁĒŚ¼ļ”¼ņĮöļ░ĢĒä░ ĒīīņØ╝ļĪ£ļ”¼ ĒÜ©ļŖźņØ┤ threonineņŚÉ ĒŖ╣ņØ┤ĒĢ©ņØä ņ”Øļ╣ÖĒĢśĻĖ░ ņ£äĒĢ£ ņŚ░ĻĄ¼ Ļ▓░Ļ│╝, group 6Ļ│╝ group 7Ļ│╝ Ļ░ÖņØ┤ threonine Ļ│╝ļ”ĮĻ│╝ C. glutamicumĻ│╝ Ļ░ÖņØĆ threonine ņāØņä▒Ļ░ĆļŖź ĻĘĀņŻ╝ņØś brothļź╝ Ļ░ÖņØ┤ ļäŻņØĆ ĻĄ░ņŚÉņä£ Ļ░Ćņן ņ£ĀņØśĒĢ£ ĒĢŁĒŚ¼ļ”¼ņĮöļ░ĢĒä░ ĒīīņØ╝ļĪ£ļ”¼ Ļ▓░Ļ│╝ļź╝ ņ¢╗ņŚłļŗż. ņØ┤ļź╝ ņóĆ ļŹö ņ”Øļ╣ÖĒĢśĻĖ░ ņ£äĒĢśņŚ¼ ņ¢╗ņ¢┤ņ¦ä brothļź╝ 1/4, 1/40 ĻĘĖļ”¼Ļ│Ā 1/100ļĪ£ ĒؼņäØĒĢ£ ņŚ░ĻĄ¼ņŚÉņä£ņØś Ļ▓░Ļ│╝ļź╝ ĒåĄĒĢ®ĒĢśņŚ¼ņä£ļÅä Ļ░ÖņØĆ Ļ▓░Ļ│╝ļź╝ ņ¢╗ņØä ņłś ņ׳ņ¢┤(Supplementary Fig. 2), ņØ┤ņāüņØś Ļ▓░Ļ│╝ļŖö threonine Ļ│╝ļ”Į ļŗ©ļÅģņØĆ ļ¼╝ļĪĀ ņØ┤ļź╝ ņāØņä▒ĒĢśļŖö ĻĘĀņŻ╝ņÖĆņØś ļ│æņÜ®ņ£╝ļĪ£ ĒĢŁĒŚ¼ļ”¼ņĮöļ░ĢĒä░ ĒÜ©ļŖźņØ┤ ņ׳ņØīņØä ņ”Øļ╣ÖĒĢĀ ņłś ņ׳ņŚłļŗż.
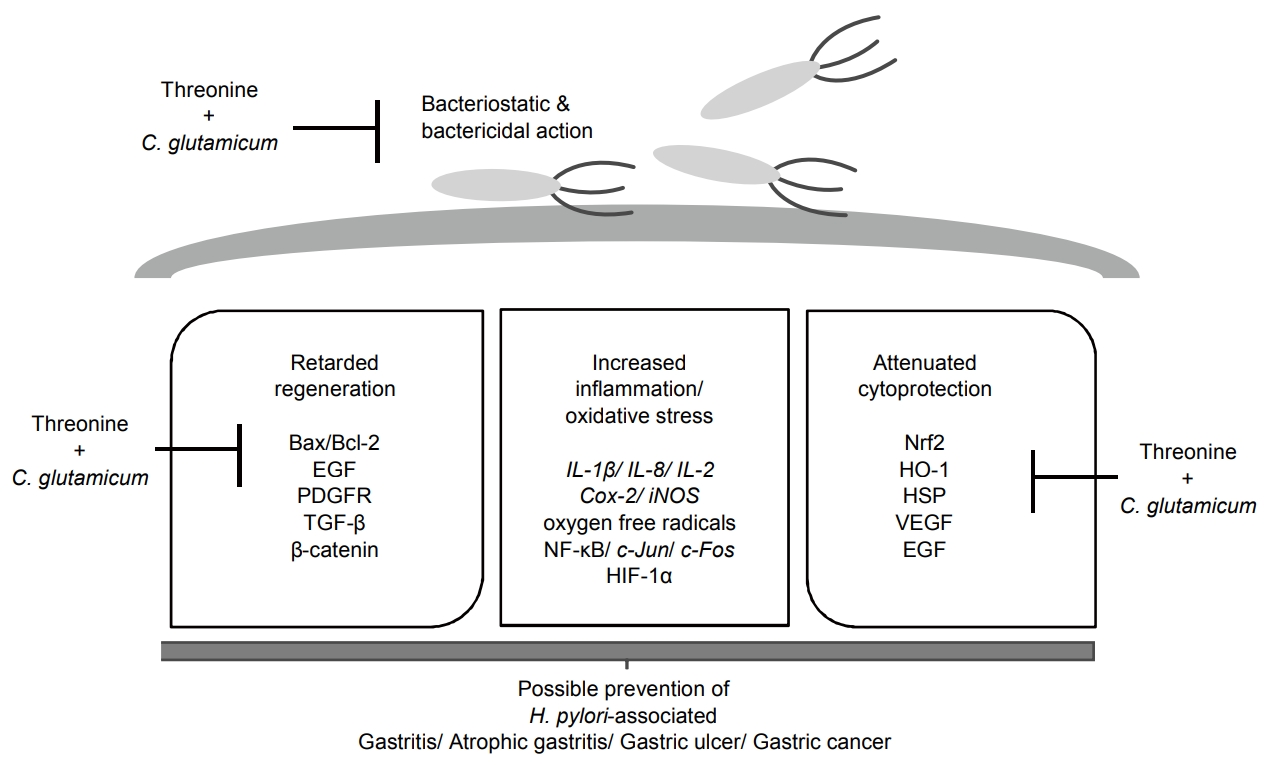
Ļ│Ā ņ░░
ņØ┤ņāüņØś ņŚ░ĻĄ¼ Ļ▓░Ļ│╝ļź╝ Fig. 7ņŚÉ Ļ░äļץĒ׳ ņĀĢļ”¼ ļ░Å ņÜöņĢĮĒĢśņśĆļŖö ļ░ö, ĒŚ¼ļ”¼ņĮöļ░ĢĒä░ ĒīīņØ╝ļĪ£ļ”¼ Ļ░ÉņŚ╝ņØĆ IL-1╬▓, IL-8, IL-2, Cox-2, iNOS, NF-╬║B ĒÖ£ņä▒, c-JunĻ│╝ c-Fos ņ”ØĻ░Ć ĻĘĖļ”¼Ļ│Ā HIF-1╬▒ņØś ņ”ØĻ░ĆņŚÉ ĻĖ░ļ░śĒĢśņŚ¼ ĻĖēņä▒/ļ¦īņä▒ ņ£äņŚ╝ņØĆ ļ¼╝ļĪĀ ļ¦īņä▒ ņ£äņČĢņä▒ ņ£äņŚ╝ņØ┤ļéś ņĀäņĢö ļ│æļ│ĆņŚÉ ĒĢ┤ļŗ╣ĒĢśļŖö ņĢöĒÖöļź╝ ņ£ĀļÅäĒĢ£ļŗż. ņØ┤ļ¤¼ĒĢ£ ļ│æĒā£ņāØļ”¼ļź╝ ņ”ØĻ░Ģņŗ£ĒéżļŖö ļśÉ ļŗżļźĖ ĻĖ░ņĀäņ£╝ļĪ£ Nrf-2, HO-1, Ļ░üņóģ growth factors ļō▒Ļ│╝ Ļ░ÖņØĆ cytoprotectionņØś ņĢĮĒÖöņŚÉ ļö░ļØ╝ ņ×äņāüņĀüņ£╝ļĪ£ļŖö ņ£ä, ņŗŁņØ┤ņ¦Ćņן ĻČżņ¢æ ļ░£ņāØ ņ”ØĻ░Ć ļ░Å ĻČżņ¢æņØś ņ╣śņ£ĀĻ░Ć ļŖ”ņ¢┤ņ¦ĆĻ│Ā, oxidative stress ĻĖēņ”ØņŚÉ ļö░ļźĖ ņäĖĒż ļ│┤ĒśĖ Ļ░Éņåī ĻĘĖļ”¼Ļ│Ā ņŗżņ¦łņĀüņØĖ apoptosisņØś ņ”ØĻ░ĆņÖĆ ņØ┤ĒøäņØś ņĀüņĀłĒĢ£ ņäĖĒż ņä▒ņן ņĪ░ņĀłņØś ņŗżĒī©ņŚÉ ļö░ļźĖ ņĢöĒÖöņØś ļŗ©ņ┤łĻ░Ć ņĀ£Ļ│ĄņØ┤ ļÉśļŖö Ļ▓āņ£╝ļĪ£ ņĢäņŻ╝ ņל ļ░ØĒśĆņĀĖ ņ׳ļŗż[1-3]. ļ│Ė ņŚ░ĻĄ¼ņŚÉņä£ļŖö ņØ┤ļ¤¼ĒĢ£ ļ░®ņ¢┤ņØĖņ×ÉņØś Ļ░ÉņåīņÖĆ Ļ░ÖņØĆ ļ░░Ļ▓ĮņŚÉ ļīĆĒĢśņŚ¼ threonine ļŗ©ļÅģ Ēł¼ņŚ¼ļŖö ļ¼╝ļĪĀ ņØ┤ļź╝ ņāØņä▒ĒĢśļŖö ĻĘĀņŻ╝ņØĖ Corynebacterium, ĒŖ╣Ē׳ C. glutamicumņØś ļ│æņÜ® Ēł¼ņŚ¼ļŖö Ļ░Ćņן ņ£ĀņÜ®ĒĢ£ ĒĢŁĒŚ¼ļ”¼ņĮöļ░ĢĒä░ ĒīīņØ╝ļĪ£ļ”¼ ĒÜ©ļŖźĻ│╝ ĒĢ©Ļ╗ś ņ£äņŚÉ ņŚ┤Ļ▒░ĒĢ£ Ļ░ÉņŚ╝ņŚÉ ļö░ļźĖ ņĀ£ļ░ś ļ│æĒā£ņāØļ”¼ĒĢÖņĀü ĻĖ░ņĀäņØä ĒÜ©ņ£©ņĀüņ£╝ļĪ£ ņĀĆĒĢ┤ĒĢśņŚ¼ņżīņØä ņ”Øļ╣ÖĒĢĀ ņłś ņ׳ņ¢┤, ņØ┤ļź╝ ņØæņÜ®ĒĢśļ®┤ threonineņØĆ ĒŚ¼ļ”¼ņĮöļ░ĢĒä░ ĒīīņØ╝ļĪ£ļ”¼ Ļ░ÉņŚ╝ņØ┤ļéś ĻĖ░ĒāĆņØś ņŚ¼ļ¤¼ Ļ░Ćņ¦Ć ņ£äņĀÉļ¦ē ņ×ÉĻĘ╣ ļ¼╝ņ¦łņØś ņ”ØĻ░Ć ņāüĒÖ®ņŚÉņä£ ņØ┤ļź╝ ĒÜ©ņ£©ņĀüņ£╝ļĪ£ ļ░®ņ¢┤Ļ░Ć Ļ░ĆļŖźĒĢ£ ņ╣śļŻīņĀ£ļĪ£ņä£ņØś Ļ░ĆļŖźņä▒ņØ┤ ņ׳ļŗżĻ│Ā ĒĢśĻ▓Āļŗż.
ĻĘĖļ¤¼ļ»ĆļĪ£ ņØ┤ļź╝ ņØĖņ▓┤ ņ¦łĒÖśņŚÉļÅä ņØæņÜ®ņØ┤ Ļ░ĆļŖźĒĢśļéś ņØ┤ļ»Ė ņØĖĻ░äņØĆ threonineļź╝ ĒĢ©ņ£ĀĒĢ£ ļ¦ÄņØĆ ņØīņŗØņØ┤ļéś ņŗØĒÆłņØä ņČ®ļČäĒ׳ ņäŁņĘ©ĒĢśĻĖ░ ļĢīļ¼ĖņŚÉ, ņØ┤ļ¤¼ĒĢ£ Ļ░ÉņŚ╝ņŚÉ ļö░ļźĖ ĻĘ╝ļ│ĖņĀüņØĖ ņśüņ¢æ Ļ│ĄĻĖēĻ│╝ Ļ░ÖņØĆ ļīĆņ▒ģņØ┤ Ļ▒░ņØś ņŚåļŖö ļÅÖļ¼╝ņØ┤ļéś Ļ░ĆņČĢņØś ņé¼ļŻīļĪ£ņØś ĒÖ£ņÜ® ļō▒ņ£╝ļĪ£ ĻĖ░ļīĆĻ░Ć ļÉśĻ▓Āļŗż[11,12].
ņ£äņןņØś ļ░®ņ¢┤ņØĖņ×É ņżæ Ļ░Ćņן ņĄ£ņĀäņäĀņŚÉ ņ׳Ļ│Ā ņżæņÜöĒĢśļ®░ pH 2, 3Ļ│╝ Ļ░ÖņØĆ Ļ░Ģņé░ņØś ņ£äņé░ņØä ņżæĒÖöņŗ£Ēé¼ ņłś ņ׳ļŖö ļ░®ņ¢┤ĻĖ░ņĀäņØ┤ ļ░öļĪ£ mucinņØś ĒĢ®ņä▒ņØĖļŹ░, HCO3-ļź╝ ļ│┤ņ£ĀĒĢśĻ│Ā pepsin ļŗ©ļ░▒ ļČäĒĢ┤ļź╝ ņ¢ĄņĀ£ņŗ£Ēé¼ ņłś ņ׳Ļ│Ā, ņØĖņ¦Ćņ¦łņØś ļČłņłśņÜ® ņä▒ņ¦łņØä ņ£Āņ¦Ćņŗ£ņ╝£ņŻ╝ļ®░, ņĀÉņĢĪ gradientļź╝ ļ¦īļōżņ¢┤ ņŻ╝ļŖö ņ£ĀņÜ®ĒĢ£ ņĀÉņĢĪņ¦łņØ┤ ļ░®ņ¢┤ĻĖ░ņĀä ņżæņŚÉ ņĢäņŻ╝ ņżæņÜöĒĢśļŗż ĒĢśĻ▓Āļŗż. ņØ┤ļ¤¼ĒĢ£ ņ£ä mucinņØĆ ņ£äĒæ£ļ®┤ ņĀÉļ¦ēņäĖĒżļéś ņ£äĻ▓ĮļČĆ ņĀÉņĢĪņäĖĒżņŚÉņä£ ņāØņä▒ļÉśļ®░, ĒÅ┤ļ”¼ļ©Ėļź╝ ĒśĢņä▒ĒĢśĻĖ░ ņ£äĒĢśņŚ¼ disulfide bridgeļĪ£ ņĀæļ¬®ļÉ£ glycoprotein subunitņ£╝ļĪ£ ĻĄ¼ņä▒ļÉśņ¢┤ ņ׳ļŖöļŹ░, ņØ┤ļĢī ļŗ╣ņćäļĪ£ ĻĄ¼ņä▒ļÉ£ ņĢäļ»ĖļģĖņé░ ļ╝łļīĆĻ░Ć ĻĄ¼ņĪ░ņāü Ļ░Ćņן ņżæņÜöĒĢśļŗż. ņØ┤ ņČĢņŚÉņä£ ļ░öļĪ£ threonineņØ┤ ņŻ╝ņÜö ĻĄ¼ņä▒ņøÉņØ┤ļ»ĆļĪ£, threonineņØś ņäŁņĘ©ļŖö ņØ┤ļ¤¼ĒĢ£ ļŗ╣ĒÖö ļ░śņØæņØś ņ”ØĻ░Ć ļ░Å ņČĢĒśĢņä▒ņ£╝ļĪ£ ņ¢æņ¦łņØś mucinņØ┤ ņāØņä▒ļÉśĻ▓ī ĒĢśļ»ĆļĪ£ ņĀäņ▓┤ņĀüņ£╝ļĪ£ ņ£ä ļ░®ņ¢┤ņØĖņ×ÉņØś ņ”ØĻ░Ćļź╝ Ļ░ĆņĀĖņśżĻ▓ī ļÉ£ļŗż. ņØ┤ļ¤¼ĒĢ£ ņČĢņØś ņ×æņÜ®ņŚÉ ļīĆĒĢ┤ņä£ļŖö ņĢäņ¦ü ĻĘ£ļ¬ģļÉśņ¦Ć ļ¬╗ĒĢśĻ│Ā ņ׳ņŚłņ£╝ļéś, ņĀĆņ×ÉļōżņØĆ ņłśņ╣© ļ░Å ĻĄ¼ņåŹ ņŖżĒŖĖļĀłņŖż ļ¬©ļŹĖņØ┤ļéś ļŗżļźĖ ĒÖöĒĢÖņĀü ņ×ÉĻĘ╣ ļ¬©ļŹĖ ņŚ░ĻĄ¼ņŚÉņä£ ņČöĻ░ĆņĀüņØĖ ļ░®ņ¢┤ņØĖņ×É ņ”ØĻ░Ģ ĻĖ░ļŖźņØä ĻĘ£ļ¬ģĒĢ£ ļ░ö ņ׳ļŗż[15,16,19-21].
ņĢłņĀĢņä▒ņØ┤ ļåÆĻ│Ā ļŹö ļ¦ÄņØĆ ĒÜ©ļŖźņØ┤ ĻĖ░ļīĆļÉśļŖö threonineņØä ņāØņä▒ Ļ░ĆļŖźĒĢ£ C. glutamicumņØĆ L-threonine ņāØņä▒ņØä ĒĢśļŖö ņĢäņŻ╝ ņżæņÜöĒĢ£ ĻĘĀņŻ╝ņØĖļŹ░, ņØ┤ļ¤¼ĒĢ£ ĻĖ░ņłĀņØĆ ņØ┤ļ»Ė 2000ļģäĻ▓ĮņŚÉ ņÖäņä▒ņØ┤ ļÉśņŚłļŗż. ņäĖĻĘĀņØ┤ ņĢäļ»ĖļģĖņé░ ņāØņä▒ņŚÉ ĒĢäņÜöĒĢ£ ĒÄ®ĒāĆņØ┤ļō£ļź╝ ņČ®ļČäĒ׳ ņäŁņĘ©ĒĢśĻ│ĀņØ┤ļź╝ ņ▓śļ”¼ĒĢ©ņ£╝ļĪ£ņŹ©, ļ│Ė ņŚ░ĻĄ¼ņŚÉņä£ ĒāĆĒéĘņØä ĒĢśļŖö L-threonine ņØ┤ņÖĖņŚÉļÅä L-lysine, putrescineĻ│╝ Ļ░ÖņØĆ ļŗżņ¢æĒĢ£ ņĢäļ»ĖļģĖņé░ņØĆ ļ¼╝ļĪĀ Ļ┤æņ╣śļŻīņŚÉ ņé¼ņÜ®ļÉśļŖö aminolevulinic acid ļō▒ļÅä ļ¦īļōż ņłś ņ׳Ļ▓ī ļÉśņŚłļŗż. ĻĘĖļ¤¼ļ»ĆļĪ£ ņØ┤ļ¤¼ĒĢ£ ĻĖ░ņłĀņØĆ ļ│Ė ņŚ░ĻĄ¼ņŚÉņä£ ņé¼ņÜ®ļÉśļŖö L-threonineņØä ņĢäņŻ╝ ļåÆņØĆ ņł£ļÅäļĪ£ ļīĆļ¤ē ņāØņé░ņØ┤ Ļ░ĆļŖźĒĢśĻ▓ī ĒĢ©ņ£╝ļĪ£ņŹ© ļ│Ė ņŚ░ĻĄ¼ņŚÉņä£ ņĀ£ņŗ£ĒĢ£ ņ£äņן ļ│┤ĒśĖ ĒÜ©ļŖźņØĆ ļ¼╝ļĪĀ ņäĖĒż ņ×¼ņāØ ļō▒ņŚÉ ļŗżņ¢æĒĢśĻ▓ī ņé¼ņÜ®ņØ┤ Ļ░ĆļŖźĒĢśĻ│Ā, ļŹö ļéśņĢäĻ░Ćņä£ ņ£äņן ņØ┤ņÖĖņØś ļŗżņ¢æĒĢ£ ņ×¼ņāØņØä ņÜöĒĢśļŖö ņ¦łļ│æņŚÉļÅä ņé¼ņÜ®ņØ┤ Ļ░ĆļŖźĒĢśĻ▓ī ļÉśņŚłļŗż[22-25].
ĒŚ¼ļ”¼ņĮöļ░ĢĒä░ Ļ░ÉņŚ╝ņŚÉ ņØśĒĢ£ ņ£äņןļ│æņØś ļ░£ņāØņØĆ Ļ░ÉņŚ╝ ņŗ£ĻĖ░ ļ░Å Ļ░ÉņŚ╝ņŚÉ ļö░ļźĖ ņłÖņŻ╝ņØś ļ░śņØæņŚÉ ļö░ļØ╝ ņĢäņŻ╝ ļŗżņ¢æĒĢ£ļŹ░, ņØ╝ļ░śņĀüņ£╝ļĪ£ļŖö ĻĖēņä▒/ļ¦īņä▒ ņ£äņŚ╝ņØä ņ£Āļ░£ĒĢ£ļŗż. ņØ┤ļĢī ņ¦Ćļéśņ╣£ ņ×ÉĻ░Ćļ®┤ņŚŁ ļ░śņØæ, ĒŖ╣Ē׳ ļ”╝ĒöäĻĄ¼ ļ░śņØæņ£╝ļĪ£ ņĀÉļ¦ēĻ┤ĆļĀ©ļ”╝ĒöäņĪ░ņ¦ü(mucosa-associated lymphoid tissue, MALT) ļ”╝ĒöäņóģņØ┤ ļ░£ņāØĒĢśļ®░, ļ░śļ®┤ņŚÉ ņ£äņé░ ļČäļ╣äņĪ░ņĀł ņןņĢĀļź╝ ņ£Āļ░£ĒĢĀ Ļ▓ĮņÜ░ ņŗŁņØ┤ņ¦Ćņן ĻČżņ¢æņØ┤ ĻĘĖļ”¼Ļ│Ā ņ¦Ćļéśņ╣£ ņŚ╝ņ”Øļ░śņØæ ņĢģĒÖö ļ░Å ņ¦ĆņåŹņ£╝ļĪ£ļŖö ņ£äņäĀ ĒīīĻ┤┤ļź╝ ņ┤łļלĒĢśļŖö Ļ▓░Ļ│╝ļĪ£ ļ¦īņä▒ ņ£äņČĢņä▒ ņ£äņŚ╝ ļ░Å ņ¦äņĀäņŚÉ ļö░ļźĖ ņ£äņĢöņØ┤ ļ░£ņāØĒĢĀ ņłś ņ׳ļŗż. ĻĘĖļ¤░ļŹ░ ņØ┤ļ¤¼ĒĢ£ ņłÖņŻ╝ ļ░śņØæņØś Ļ│ĄĒåĄņĀÉņØĆ Ļ░ÉņŚ╝ ļ░Å ņ¦æļØĮĒÖö Ļ▓░Ļ│╝ļĪ£ ņé░ĒÖöņĀü ņåÉņāü, ļÅÖļ░śļÉśļŖö ņ¦Ćļéśņ╣£ ņŚ╝ņ”Ø ļ░śņØæ ĻĘĖļ”¼Ļ│Ā ņØ┤ņØś ļ│æņØĖņØ┤ ņ¦ĆņåŹļÉśļŖö ņłÖņŻ╝ņØś ņäĖĒż ļ│┤ĒśĖ ņĢĮĒÖö ļō▒ņØ┤ ĻĖ░ļ│ĖņĀüņ£╝ļĪ£ ļ░£ņāØĒĢ£ļŗżļŖö Ļ▓āņØ┤ļŗż. ņØ┤ļ¤¼ĒĢ£ ļ░░Ļ▓Įņ£╝ļĪ£ ņ×äņāüņŚÉņä£ļŖö ņåīĒÖöņä▒ ĻČżņ¢æ ļ░£ņāØ ļ░Å ņ×¼ļ░£, MALT ļ”╝ĒöäņóģĻ│╝ Ļ░ÖņØĆ ņ¦Ćļéśņ╣£ ļ”╝ĒöäĻĄ¼ ļ░śņØæ ĻĘĖļ”¼Ļ│Ā ņ£äņĢö ļ░£ņāØĻ│╝ ņŚ░Ļ┤ĆļÉ£ ņ×¼ļ░£ ļ░®ņ¦Ć ļō▒ņØä ņ£äĒĢ£ ļ¬®Ēæ£ļĪ£ļ¦ī ņĀ£ĻĘĀ ņĀüņØæņ”ØņØ┤ ņĀ£ņĀĢļÉśņ¢┤ ņ׳ņ¦Ćļ¦ī, ļ│æĒā£ņāØļ”¼ĒĢÖņĀü ņĖĪļ®┤ņŚÉņä£ļŖö Ļ░ÉņŚ╝ ļ░Å ņŗ¼ĒĢ£ ņŚ╝ņ”ØĻ│╝ ļ®┤ņŚŁ ļ│ĆĒÖöņŚÉ ļö░ļźĖ ņ┤łĻĖ░ ļ░śņØæņŚÉ ļīĆĒĢ£ ņĀüņĀłĒĢ£ ņĀ£ņ¢┤Ļ░Ć ņĢäņŻ╝ ņżæņÜöĒĢśļŗż ĒĢśĻ▓Āļŗż. ĻĘĖļ¤¼ļ»ĆļĪ£ ņØ┤ļ¤¼ĒĢ£ ļ░░Ļ▓Į ĒĢśņŚÉ ņé¼ļ×īņØĆ ļ¼╝ļĪĀ ņ×ÉĻĘ╣ ļ¼╝ņ¦łņŚÉ ļīĆĒĢ£ ņ¦Ćļéśņ╣£ ņ¦łĒÖś ļ░śņØæņ£╝ļĪ£ ĒÅÉņé¼ļź╝ ĒĢśĻ▒░ļéś ņ£äņן ņ¦łĒÖśņØ┤ ļ│æļ░£ĒĢśĻ│Ā, ĒŖ╣Ē׳ ņŖżĒŖĖļĀłņŖżļź╝ ņØ┤Ļ▓©ļé┤ļŖö ĻĖ░ņĀäņØ┤ļéś ĒÖśĻ▓ĮņØä ļŖźļÅÖņĀüņ£╝ļĪ£ ļīĆņ▓śĒĢĀ ņłś ņŚåļŖö ļÅÖļ¼╝ļōżņŚÉĻ▓īļŖö ņé¼ļŻīņŚÉ ĒżĒĢ©ņØä ņŗ£ĒéżĻ▒░ļéś ļ│äļÅäļĪ£ Ļ│ĄĻĖēĒĢśļŖö ļ░®ļ▓Ģņ£╝ļĪ£ threonineņØä ņäĀņĀ£ņĀü Ēł¼ņŚ¼(preemptive administration)ĒĢśļŖö Ļ▓āņØ┤ ņĢäņŻ╝ ņĀüņĀłĒĢĀ Ļ▓āņØ┤ļŗż[26,27].
Threonine Ēł¼ņŚ¼ņØś Ļ▓░Ļ│╝ļĪ£ ņĢäņŻ╝ ņ£ĀņÜ®ĒĢ£ ĒĢŁņŚ╝ņ”Ø ņ×æņÜ®, ĒĢŁņé░ĒÖö ņ×æņÜ®, ĒĢŁļ│ĆņØ┤ ņ×æņÜ® Ļ▓░Ļ│╝ļź╝ ļ│Ė ņŚ░ĻĄ¼ņŚÉņä£ ĻĘ£ļ¬ģĒĢśņśĆĻ│Ā, ņØ┤ņĀäņØś ņŚ░ĻĄ¼ņŚÉņä£ļÅä ņŖżĒŖĖļĀłņŖż ņŚ░Ļ┤Ć ņ£ä ņåÉņāü ļ¬©ļŹĖņŚÉņä£ļÅä ņĀÉņĢĪ ņāØņä▒ ņ”ØĻ░ĆļŖö ļ¼╝ļĪĀņØ┤Ļ│Ā ĒĢŁņé░ĒÖö, ĒĢŁņäĖĒż ņé¼ļ®Ė ĻĘĖļ”¼Ļ│Ā ņäĖĒż ņ×¼ņāØ ņ┤ēņ¦ä ļō▒ņØś Ļ│ĄĒåĄļÉ£ ĒÜ©Ļ│╝ļź╝ ņØ┤ļ»Ė Ļ┤Ćņ░░ĒĢśņŚ¼[16] threonineņØś Ēł¼ņŚ¼Ļ░Ć ļÅÖļ¼╝ņØś ņ£ä ņåÉņāü ņśłļ░®ņØ┤ļéś ņ╣śņ£ĀņŚÉ ņé¼ņÜ®ņØ┤ Ļ░ĆļŖźĒĢĀ Ļ▓āņ£╝ļĪ£ ņé¼ļŻīļÉśļ®░, ļŹö ļéśņĢäĻ░Ć ņé¼ļ×īņŚÉņä£ļÅä ņØæņÜ®ņØ┤ ĻĖ░ļīĆļÉ£ļŗżĻ│Ā ĒĢśĻ▓Āļŗż. ņ£äņĀÉņĢĪņØĆ Ēæ£ļ®┤ ņ£äņĀÉļ¦ēņäĖĒżņÖĆ ņ£äĻ▓ĮļČĆ ņĀÉņĢĪļČäļ╣äņäĖĒżņŚÉņä£ ĒÖ£ļ░£ĒĢśĻ▓ī ļČäļ╣äļÉśņ¢┤ ņĄ£ņĀäņäĀņØś ņ£ä ļ░®ņ¢┤ņØĖņ×ÉļĪ£ ņ×æņÜ®ņØä ĒĢśļŖöļŹ░, ĻĘĖ ņāØņä▒ Ļ│╝ņĀĢņØä ņé┤ĒÄ┤ļ│┤ļ®┤ ļ░öļĪ£ ļ│Ė ņŚ░ĻĄ¼ņŚÉ ņé¼ņÜ®ļÉ£ threonine ņĢäļ»ĖļģĖņé░ņØä ĻĖ░ļ░śņ£╝ļĪ£ ļ”¼ļ│┤ņå£ņŚÉņä£ ļŗ╣ļŗ©ļ░▒ņØ┤ ņāØņä▒ļÉśņ¢┤ glycoside sugar chainņØä ņåīĒżņ▓┤ņŚÉņä£ ĻĄ¼ņä▒ĒĢśņŚ¼, oligomerization ĻĖ░ņĀäņØä ĒåĄĒĢśņŚ¼ ņä▒ņłÖļÉśļ®┤ņä£ ļČäļ╣äĻ░Ć ņ¦äĒ¢ēņØ┤ ļÉ£ļŗż.
ņĀ£ĻĘĀ ņØ┤ņÖĖņØś ņĀæĻĘ╝ ļ░®ļ▓Ģņ£╝ļĪ£ ĒŚ¼ļ”¼ņĮöļ░ĢĒä░ ĒīīņØ╝ļĪ£ļ”¼ņŚÉ ņØśĒĢ£ ņŚ¼ļ¤¼ Ļ░Ćņ¦ĆņØś ņäĖĒż ļÅģņä▒ņØä ņĢĮĒÖöņŗ£ĒéżĻ▒░ļéś ņäĖĒż ļ│Ćņä▒ņØä ņ£ĀļÅäĒĢśļŖö Ļ░üņóģ ĻĖ░ņĀäņØä ņĀ£ņ¢┤ĒĢĀ ņłś ņ׳ļŖö ņŗØĒÆłņØ┤ļéś ļ╣äņĀ£ĻĘĀņØś ļ░®ļ▓Ģņ£╝ļĪ£ļŖö ļŗżņ¢æĒĢ£ ņŗØļ¼╝ĒÖöĒĢ®ļ¼╝(phytochemicals)ņØ┤ ĒĢ©ņ£ĀļÉ£ ņŗØĒÆłņØ┤ļéś ļČäļ”¼ļÉ£ ņŗØĒÆłĒÖöĒĢ®ļ¼╝ņØä ņØ┤ņÜ®ĒĢĀ ņłś ņ׳ņ¢┤, ņĀĆņ×ÉļōżņØĆ Ļ│╝Ļ▒░ 20ņŚ¼ļģä ļÅÖņĢł ĒÖŹņé╝, Ļ░Éņ┤ł, ļģ╣ņ░©, Ļ░ĢĒÖ®, ņæź ņČöņČ£ļ¼╝ ļō▒ ņĢäņŻ╝ ļŗżņ¢æĒĢ£ ņĀæĻĘ╝ņØä ĒåĄĒĢ£ ņŚ░ĻĄ¼ļōżņØä ļ░£Ēæ£ĒĢ£ ļ░ö ņ׳ļŖöļŹ░, ņØ┤ļōżņØś Ļ│ĄĒåĄņĀüņØĖ ĒŖ╣ņ¦ĢņØĆ ņäĖĒż ļ¬©ļŹĖņØĆ ļ¼╝ļĪĀ ņ¦łĒÖś ļ¬©ļŹĖ ĻĖ░ļ░śņØś ņ£ĀĒÜ©ņä▒ņŚÉņä£ļÅä ņĢäņŻ╝ ņÜ░ņłśĒĢ£ Ļ▓░Ļ│╝ļź╝ ņ░ŠņØä ņłś ņ׳ņŚłļŗż[28,29]. ĻĘĖļĀćņ¦Ćļ¦ī ņØ┤ļ¤¼ĒĢ£ ņĀæĻĘ╝ņØś ĒĢ£Ļ│äļĪ£ļŖö ņĢäņŻ╝ ņČ®ļČäĒĢ£ ĻĘ╝Ļ▒░ņżæņŗ¼ņØśĒĢÖņĀü Ļ▓░Ļ│╝Ļ░Ć ļČĆņĪ▒ĒĢśļŗżļŖö Ļ▓āĻ│╝ ņØ┤ļōż ļ¼╝ņ¦łļōżņØĆ ļÅģņä▒ĒĢÖņŚÉņä£ ņĀ£ĻĖ░ļÉśļŖö hormesisņØś ņøÉņ╣ÖņØä ļö░ļźĖļŗżļŖö Ļ▓āņØ┤ļŗż. ņ”ē, ņĢĮļ¼╝Ļ│╝ Ļ░ÖņØ┤ ņĢäņŻ╝ ĒŖ╣ņĀĢĒĢ£ ļåŹļÅäļéś Ēł¼ņŚ¼ ņŗ£ņŚÉļŖö ĒÜ©ļŖźņØä ļ│┤ņØ┤ĻĖ░ļŖö ĒĢśļéś ņÜ®ļ¤ē ņØśņĪ┤ņĀüņØ┤ņ¦Ć ņĢŖņĢä ņ¢┤ļ¢ż Ļ▓ĮņÜ░ņŚÉļŖö Ļ│ĀņÜ®ļ¤ēņŚÉņä£ ņśżĒ׳ļĀż ļé«ņØĆ ĒÜ©Ļ│╝ļéś ļČĆņ×æņÜ®ņØä ļ│┤ņØĖļŗżļŖö ņĀÉĻ│╝ ņØ┤ļōżņØś ĒÜ©ļŖźņØä ĻĘ╣ļīĆĒÖöĒĢśĻĖ░ ņ£äĒĢ£ ņĀüņĀł ņÜ®ļ¤ē ļ░Å Ēł¼ņŚ¼ ļ░®ļ▓Ģ ļō▒ņŚÉ ļīĆĒĢ£ ĻĄ¼ņ▓┤ņĀüņØĖ Ļ▓░Ļ│╝Ļ░Ć ņŚåņ£╝ļ»ĆļĪ£ ņ×äņāüņŗ£ĒŚśņØś ņłśņżĆļÅä ļé«ņ¦Ćļ¦ī ņØ╝Ļ┤Ćņä▒ņØ┤ ņ׳ļŖö Ļ▓░Ļ│╝ļź╝ ņĀ£ņŗ£ĒĢĀ ņłś ņŚåļŗżļŖö Ļ▓āņØ┤ļŗż. ĻĘĖļ¤░ļŹ░ ļ░śĒĢśņŚ¼ ļ│Ė ņŚ░ĻĄ¼ņŚÉ ņé¼ņÜ®ļÉ£ threonineņØś Ļ▓ĮņÜ░ļŖö ņØ┤ļ¤¼ĒĢ£ ļŗ©ņĀÉņØä ļ¬©ļæÉ ļČłņŗØņŗ£Ēé¼ ņłś ņ׳ņŚłĻ│Ā, ļ│Ė ņŚ░ĻĄ¼ņŚÉņä£ ņĀ£ņŗ£ĒĢ£ Ļ▓āĻ│╝ Ļ░ÖņØĆ ĒŚ¼ļ”¼ņĮöļ░ĢĒä░ ĒīīņØ╝ļĪ£ļ”¼ Ļ░ÉņŚ╝ņŚÉ ļö░ļźĖ ļ│æĒā£ņāØļ”¼ļź╝ ļ¬©ļæÉ ņ£ĀĒÜ©ĒĢśĻ▓ī ņĪ░ņĀłņØ┤ Ļ░ĆļŖźĒĢśļ®░, ņŚ¼ļ¤¼ ņĪ░Ļ▒┤ņŚÉņä£ ņØ╝ņ╣śļÉ£ Ļ▓░Ļ│╝Ļ░Ć ļéśņś©ļŗżļŖö ņןņĀÉĻ│╝ ņĢłņĀĢņä▒ņØ┤ ļåÆņØĆ ņĢäļ»ĖļģĖņé░ņØ┤ļ»ĆļĪ£ Ēśäņ×¼ļĪ£ļŖö ĒŖ╣ņØ┤ĒĢ£ ļīĆņ▒ģņØ┤ ņŚåļŖö ļÅÖļ¼╝ņØ┤ļéś Ļ░ĆņČĢņØś ņé¼ļŻīļĪ£ ņČ®ļČäĒ׳ Ļ░Ćņ╣śĻ░Ć ļåÆņØä ņłś ņ׳ļŗż.
ņØ┤ņāüņØś Ļ▓░Ļ│╝ļź╝ ņÜöņĢĮĒĢśļ®┤ ĒŚ¼ļ”¼ņĮöļ░ĢĒä░ ĒīīņØ╝ļĪ£ļ”¼ļź╝ ļ¬©ļæÉ ņĀ£Ļ▒░ĒĢśĻ▒░ļéś ņØ┤ļź╝ ĒÜ©ņ£©ņĀüņ£╝ļĪ£ ņ¢ĄņĢĢĒĢśĻĖ░ ņ£äĒĢ£ ņĀ£ļ░ś ļģĖļĀźņØĆ ĒŚ¼ļ”¼ņĮöļ░ĢĒä░ ĒīīņØ╝ļĪ£ļ”¼ Ļ░ÉņŚ╝ņŚÉ ņØśĒĢ£ ĻĖēņä▒/ļ¦īņä▒ ņ£äņŚ╝ņØś ņ╣śļŻīļŖö ļ¼╝ļĪĀ, ņåīĒÖöņä▒ ĻČżņ¢æ, MALT ļ”╝ĒöäņóģņØ┤ļéś ņ£äņäĀņĢöņØś ņśłļ░®, ņĄ£ĻĘ╝ņŚÉļŖö Ļ░üņóģ ņ£äņן ņÖĖņØś ņŚ¼ļ¤¼ ņ¦łĒÖśļōżņØś ņøÉņØĖņ£╝ļĪ£ ĻĘ£ļ¬ģļÉśĻ│Ā ņ׳ņ¢┤ ņĀÉņ░© ņØ┤ ĻĘĀņŻ╝ņŚÉ ņØśĒĢ£ ņŚ░Ļ┤Ć ņ¦łĒÖśņØś ņ╣śļŻīļĪ£ ņĀüņØæņ”ØņØ┤ ĒÖĢļīĆļÉśĻ│Ā ņ׳ļŗż. ņØ┤ļ¤¼ĒĢ£ ņ£äņן ņÖĖ ņ¦łĒÖśņØś Ļ░ĆļŖźĒĢ£ ļ│æņØĖņ£╝ļĪ£ ĒŚ¼ļ”¼ņĮöļ░ĢĒä░ ĒīīņØ╝ļĪ£ļ”¼ exosomeņŚÉ ņØśĒĢ£ Ļ░ĆļŖźņä▒Ļ│╝ molecular mimicryņŚÉ ņØśĒĢ£ ņ×ÉĻ░Ć ļ░śņØæņ£╝ļĪ£ ļ░ØĒśĆņĀĖ ņ׳ņ£╝ļéś ņĢäņ¦üĻ╣īņ¦Ć ņé¼ļ×īņŚÉņä£ļÅä ĻĘ£ļ¬ģļÉśņ¦Ć ļ¬╗ĒĢ£ ņāüĒÖ®ņŚÉņä£ ļÅÖļ¼╝ņØ┤ļéś Ļ░ĆņČĢļōżņŚÉņä£ ļ©╝ņĀĆ Ļ│ĀļĀżļÉśņ¢┤ņĢ╝ ĒĢĀ ņé¼ĒĢŁņØĆ ņĢäļŗłļŗż. ņØ┤ļ¤¼ĒĢ£ ļ░░Ļ▓ĮņŚÉņä£ ļ│Ė ņŚ░ĻĄ¼ņ¦äļōżņØ┤ ĻĘ£ļ¬ģĒĢ£ threonine Ļ│╝ļ”ĮņØĆ Ē¢źĒøä ļÅÖļ¼╝ņØś ņé¼ļŻīņŚÉ ņé¼ņÜ®ļÉśĻ▒░ļéś ļŗ©ļÅģņ£╝ļĪ£ ļ│┤ņČ®ĒĢ┤ņżä ņłś ņ׳ļŖö ņĢäņŻ╝ ņ£ĀņÜ®ĒĢ£ ņä▒ļČäņ×äņØä ĻĘ£ļ¬ģĒĢśņśĆĻ│Ā ļŗżĒ¢ēĒ׳ļÅä ļŗ©ļ░▒ņ¦ł ņ×¼ņĪ░ĒĢ® ļ░®ļ▓Ģ ļō▒ņØś ĒśäņĀĆĒĢ£ ļ░£ņĀäņ£╝ļĪ£ ļ│Ė ņŚ░ĻĄ¼ņÖĆ Ļ░ÖņØ┤ C. glutamicumĻ│╝ Ļ░ÖņØĆ ĻĘĀņŻ╝ņØś ĒÖ£ņÜ®ņ£╝ļĪ£ ĒŚ¼ļ”¼ņĮöļ░ĢĒä░ ĒīīņØ╝ļĪ£ļ”¼ļŖö ļ¼╝ļĪĀ ņØ┤ņÖĖņØś ļŗżņ¢æĒĢ£ ņ£äņן ņ×ÉĻĘ╣ ļ¼╝ņ¦łļōżņŚÉ ļīĆĒĢŁĒĢśļŖö ņĢäņŻ╝ ņóŗņØĆ ņ╣śļŻīļ▓ĢņØ╝ ņłś ņ׳ņØīņØä ņ×ģņ”ØĒĢĀ ņłś ņ׳ņŚłļŗż.